|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Работа №5 Определение скорости проводимости Виртуальный практикум «LupraFisim»
Цель: измерить скорость проводимости нерва, с использованием следующих типов нервов: - тонкий миелинизированный нерв лягушки; - немиелинизированный нерв крысы; - толстый миелинизированный нерв крысы. Принцип действия: воздействию электрического раздражителя подвергаются нервы разного типа, и определяется скорость их проводимости: с помощью двух электродов, размещенных на известном расстоянии от электрода-раздражителя, замеряется потенциал действия. Так как расстояние известно, то, засекая время, можно вычислить скорость проводимости. Ход работы. 1. Воздействуйте электрическим стимулом на седалищный нерв лягушки, и узнайте время, понадобившееся для того, чтобы ПД распространился на заранее определенное расстояние; определите скорость проводимости для этого типа нерва; 2. Воздействуйте электрическим стимулом на не имеющий на миелиновой оболочки нерв крысы, узнайте время, понадобившееся для того, чтобы ПД распространился на заранее определенное расстояние; определите скорость проводимости для этого типа нерва; 3. Воздействию электрическим стимулом на покрытый миелиновой оболочкой нерв крысы, узнайте время, понадобившееся для того, что ПД распространился на заранее определенное расстояние; определите скорость проводимости для этого типа нерва; 4. Сделайте вывод: как наличие или отсутствие миелинового слоя влияет на скорость проводимости нерва?
РЕЗУЛЬТАТЫ: Пороговая сила тока увеличивается при уменьшении крутизны его нарастания, а при некоторой минимальной крутизне ответы на раздражение исчезают. Это явление обозначают термином «аккомодация». Уменьшение последней приводит к повышению критического уровня деполяризации (примерно на 20 % от исходной величины) и снижению амплитуды потенциалов действия. При снижении крутизны до некоторого минимального уровня («минимальный градиент», или «критический наклон») потенциал действия не возникает. Величина этого «минимального градиента», выраженного в единицах реобаза в секунду, принята в качестве меры скорости аккомодации. В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей, развивающиеся вовремя медленно нарастающей деполяризации мембраны. Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных нервных волокон скорость аккомодации, как правило, значительно выше, чем у чувствительных волокон.
ВЫВОД: Нервные волокна обладают способностью отвечать несколькими потенциалами действия на включение постоянного деполяризующего тока. Эта способность к повторным ответам хорошо выражена у сенсорных волокон, для которых длительная деполяризация мембраны рецептора является естественным раздражителем. Как правило, пороговая сила тока, необходимая для возникновения повторных ответов, выше, чем для инициации одиночного потенциала действия. Увеличение силы деполяризующего тока до определенной величины обусловливает возрастание частоты импульсов и увеличение их числа (1—4). Однако при дальнейшем повышении силы тока частота импульсов уменьшается и в конечном итоге возникает только одиночный потенциал действия (5—8). При постоянной силе тока длина меж импульсных интервалов в повторном ответе постепенно увеличивается. Это явление получило название адаптации. В его основе лежит медленное повышение калиевой проводимости мембраны, связанное с активацией особых медленных калиевых каналов. Эти каналы найдены как в нервных волокнах, так и в нервных клетках, у которых способность к повторным ответам и явление адаптации, как правило, хорошо выражены.
Занятие №4. Морфофункциональная характеристика мышечных клеток. Классификация мышечных клеток, роль в организме. Механизм мышечного сокращения.
Домашнее задание: 1. Дать классификацию мышц. Укажите локализацию этих мышц. Значение различных видов мышц в организме. Классификация мышц по строению: 1. скелетные 2. гладкие 3. сердечная
Поперечно-полосатые мышцы делят на: 1. скелетные 2. сердечная
Перечислите функции скелетных мышц: Скелетные мышцы обеспечивают: 1. возбудимость 2. растяжимость 3. эластичность 4. пластичность
2. Нарисовать саркомер, указать его основные элементы
Схема саркомера сердечной мышцы и ионных потоков.
Этапы мышечного сокращения: 1. увеличение внутриклеточной концентрации ионов кальция 2. Caсвязывается с калмодулином (CaM) 3. CaMактивирует киназу легких цепей миозина (MLCK) 4. MLCKфосфорилирует легкие цепи миозиновых головок и увеличивает активность миозин АТРазы 5. происходит образование поперечных мостиков и скольжение миозина по актину
Роль кальция в сокращении: Данные о роли ионов кальция в сократительной активности мышц накапливались довольно медленно. Кальций активен в саркоплазме при такой низкой (10-6 М и менее) концентрации, что до открытия кальцийхелатных реагентов, например ЭДТА и ЭГТА, ее невозможно было поддерживать в экспериментальных растворах. Дело в том, что даже в бидистиллированной воде концентрация ионов кальция превышает 10-6 М. Самые первые доказательства физиологической роли Са2+ представлены в работах Рингера и Бакстона. Авторы обнаружили, что изолированное сердце лягушки прекращает сокращения при отсутствии кальция в омывающем растворе. Так появились раствор Рингера и другие физиологические солевые растворы. Камада и Киносита, а затем Хейлбрун и Вертинский проверяли участие Са2+ в регуляции мышечного сокращения путем введения разных катионов внутрь мышечных волокон. Из всех изученных ионов только кальций вызывал сокращение при концентрациях, соизмеримых с концентрациями Са2+ обычно наблюдаемыми в живой ткани. Впоследствии было обнаружено, что скелетная мышца не сокращается в ответ на деполяризацию мембраны, если исчерпаны запасы кальция во внутренних депо, а подвергнутые предварительной экстракции препараты волокон скелетной мышцы не сокращаются при добавлении АТФ, если отсутствует Са2+. Количественная зависимость между концентрацией свободного Са2+ в саркоплазме и силой мышечного сокращения была установлена сравнительно недавно. Для проведения анализа удаляли поверхностную мембрану и оголенные миофибриллы обрабатывали растворами кальция различной концентрации. Сила возрастает от нуля при концентрации кальция около 10-8 М до максимального значения при концентрации кальция около 5х10-6 М. Данная зависимость между силой и концентрацией Са2+ аналогична зависимости между АТФазной активностью (скоростью гидролиза АТФ) гомогенизированных миофибрилл и концентрацией Са2+. Такое совпадение характеристик наводило на мысль, что Са2+ служит кофактором АТФазной активности миозина. Но оказалось, что это не так. АТФазная активность чистого раствора миозина довольно низкая, но сильно возрастает при добавлении очищенного актина. Это указывает на то, что АТФазный центр миозина активируется при связывании миозина с актином. В интактной мышце активация АТФазного центра миозина осуществляется при присоединении поперечного мостика к активному филаменту. Эксперименты, проведенные в лаборатории Эбаши, показали, что тропонин и тропомиозин, лежащие вдоль актиновой спирали, препятствуют присоединению миозиновых поперечных мостиков к актину. Тропонин – единственный белок в актиновых и миозиновыхфиламентах поперечнополосатых мышц позвоночных животных, имеющий высокое химическое сродство к Са2+. Каждый тропониновый комплекс связывает четыре иона кальция. Тропониновые комплексы расположены вдоль актиновогофиламента через каждые 40 нм, прикрепляясь одновременно к актиновомуфиламенту и молекуле тропомиозина. В состоянии покоя положение тропомиозина конформационно препятствует соединению головок миозина с актиновымфиламентом. Связывая Са2+, тропонин претерпевает конформационные изменения, в результате чего молекула тропомиозина смещается и освобождает дорогу миозиновым поперечным мостикам для прикрепления к актиновым центрам. Следовательно, присоединение Са2+ к тропонину устраняет постоянно существующее препятствие для взаимодействия поперечных мостиков с актином. Из результатов экспериментов, сделан вывод, что ингибирование присоединения мостиков снимается при концентрации свободного Са2+ свыше 10-7 М.
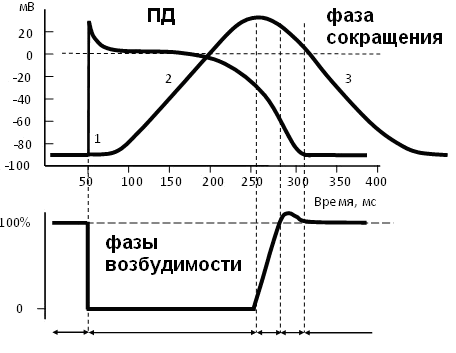
3. Изобразите синхронные графики потенциала действия, динамики возбудимости и одиночного мышечного сокращения.
4. Указать виды мышечного сокращения: 1. одиночное мышечное сокращение 2. тетаническое мышечное сокращение (тетанус) 3. тоническое мышечное сокращение
|
Последнее изменение этой страницы: 2019-04-11; Просмотров: 2453; Нарушение авторского права страницы