|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
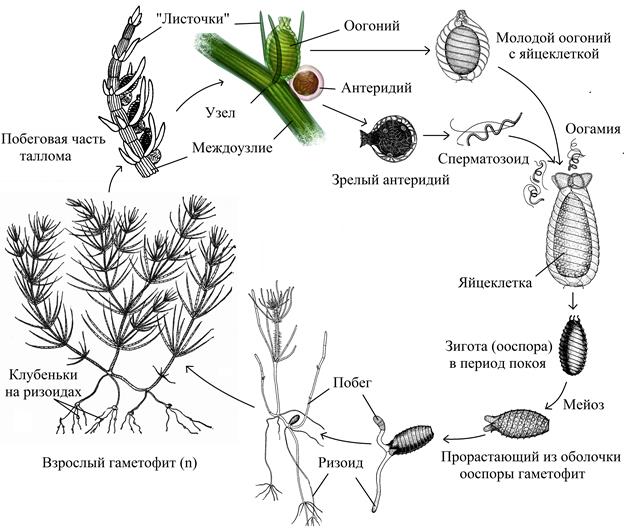
Жизненный цикл развития зелёной водоросли хара
Харовые водоросли, или лучицы (Charophyceae) – класс некогда обширной группы древних растений, которые объединяют в себе признаки водорослей и высших растений. Название происходит от др.-греч.χ ᾰ ρ ά – радость, красота. Всего известно не более 700 видов харовых. В цикле развития харовых водорослей доминирует обоеполый (у некоторых видов раздельнополый) гаметофит (n). Тело гаметофитов представляет собою много клеточный таллом высотой 20–30 (до 100–200) см. Таллом имеет метамерное строение, состоит из осевой части, на которой мутовками расположены веточки (боковые побеги). Прикреплён таллом к грунту специальными выростами – ризоидами. Рост осевой части таллома неограничен, а боковые побеги обладают предельным ростом. Каждое междоузлие метамера – это одна многоядерная гигантская, длиной до нескольких сантиметров, вытянутая клетка, неспособная к делению (у некоторых харофитов она покрыта еще корой – вторым слоем длинных клеток), тогда как каждый узел состоит из собранных в диск нескольких мелких одноядерных клеток, дифференцирующихся в процессе деления и образующих боковые побеги таллома первого – третьего порядков. Оболочки клеток иногда обызвестленные. Хлоропласты зелёные, содержат хлорофиллы a и b, из дополнительных пигментов – ликопин. Запасное вещество – крахмал.
Рис. 39. Жизненный цикл зелёной водоросли хара
Для харовых водорослей характерно вегетативное и половое размножение. Вегетативное размножение осуществляется посредством специальных клубеньков на ризоидах или звездообразных скоплений клеток (клубеньков) на нижних узлах, которые дают начало новому таллому. При половом размножении хары в узлах метамеров образуются половые органы. Женский орган – оогоний, овальный, длинной до 1 мм, состоит из яйцеклетки, которую снаружи спирально оклужают пять узких клеток. Крепится к таллому оогоний одноклеточной ножкой, сверху имеет коронку из пяти или десяти коротких клеточек. Антеридии – мужские гаметангии, находятся в узлах под женскими оогониями. Имеют шаровидную форму диаметром до 0, 5 мм, образованные восемью плоскими, скрепленными краями клетками с отходящими внутрь отростками, на которых сложным путем возникает множество мужских половых клеток. После формирования сперматозоиды выходят в воду, подплывают к коронке и ввинчиваются в оогоний. В результате кариогамии образуется покоящаяся зигота (ооспора). Зрелые зиготы могут покрываться слоем извести и накапливают запасные вещества (крахмал, липиды). Через время зиготы отделяется от материнского таллома, попадают на грунт. После периода покоя ядро зиготы делится мейозом образуя четыре гаплоидные ядра. Одно из гаплоидных ядер отделяется от остальных, в оболочке зиготы (в верхней части ооспоры), клеточной стенкой, образуя центральную линзообразную клетку. Трёхядерная клетка ооспоры выполняет питающую функцию. Центральная клетка делится, формируя большую побеговую клетку и маленькую ризоидную клетку, которые делясь развиваются соответственно в побег и ризоид нового таллома.
ГРИБЫ (Fungi или Mycota) – царство живой природы, объединяющее эукариотические организмы, сочетающие в себе некоторые признаки как растений, так и животных. Грибы изучает наука микология, которая считается разделом ботаники, поскольку ранее грибы относили к царству растений. У множества клеток грибов имеется клеточная стенка, отсутствует она лишь у зооспор и вегетативных клеток некоторых примитивных грибов. На 80–90 % она состоит из азотистых и безазотистых полисахаридов, у большинства основным полисахаридом является хитин, у оомицетов – целлюлоза. Также в состав клеточной стенки входят белки, липиды и полифосфаты. Внутри находится протопласт, окружённый цитоплазматической мембраной. Протопласт имеет строение типичное для эукариот. Есть запасающие вакуоли, содержащие волютин, липиды, гликоген, жирные кислоты (в основном ненасыщенные) и другие вещества. Ядер одно или несколько. У различных групп преобладают различные стадии по плоидности. Основа тела грибов – мицелий (грибница) – система тонких ветвящихся нитей – гиф. Грибница обычно имеет большую общую поверхность, так как через неё осмотическим путём всасывается пища. У низших грибов мицелий не имеет клеточных перегородок, то есть является синцитием. Гифы растут апикально и обильно ветвятся. При образовании органов спороношения, а иногда и вегетативных структур плотно переплетаются, образуя ложную ткань плектенхиму, иногда она может дифференцироваться на слои с различными функциями, обычно напоминает паренхиму, но в отличие от неё образуется не делением клеток, а переплетением гиф. Параллельное сплетение гиф образует мицелиальные тяжи, иногда достигающие больших размеров и называемые тогда ризоморфами (опёнок, домовый гриб). Особые видоизменения мицелия, служащие для перенесения тяжёлых условий, называются склероциями, из них развивается новый мицелий или органы плодоношения. Все грибы являются гетеротрофными организмами. Минеральные и органические вещества грибы усваивают из окружающей среды. В зависимости от потребности в веществах, грибы заселяют определённые субстраты. Для грибов характерно внешнее пищеварение, то есть сначала в окружающую среду, содержащую органические вещества, выделяются ферменты, расщепляющие вне организма полимеры до легкоусваиваемых мономеров, которые всасываются в цитоплазму. Некоторые грибы способны выделять все основные типы пищеварительных ферментов – протеазы, расщепляющие белки; липазы, расщепляющие жиры; карбогидразы, расщепляющие полисахариды, поэтому они способны поселяться практически на любом субстрате. Другие грибы выделяют лишь определённые классы ферментов и заселяют субстрат, содержащий соответствующие вещества. Размножаются грибы: вегетативно – частями мицелия; бесполым путём – эндогенными спорами (образованными у низших грибов в спорангиях) и экзогенными спорами – конидиями (сегментами специальных гиф – конидиеносцев); половым путём – зиготами (характерно для низших грибов). Для большинства грибов характерен половой процесс.
Домен Эукариоты, или Ядерные (Eucaryota) Царство Грибы (Fungi или Mycota) Подцарство Грибоподобные организмы Отдел Миксомицеты (Myxomycota) Отдел Плазмодиофоровые (Plasmodiophoromycota) Подцарство Низшие грибы Отдел Оомицеты (Oomycota) Отдел Лабиринтуловые (Labyrinthulomycota) Отдел Гифохитриевые (Hyphochytriomycota) Отдел Хитридиомицеты (Chytridiomycota) Отдел Зигомицеты (Zygomycota) Подцарство Высшие грибы Отдел Аскомицеты (Ascomycota) Отдел Базидиомицеты(Basidiomycota) Отдел Дейтеромицеты (Deuteromycota)
Зигомицеты (Zygomycota) – отдел грибов, объединяющий 10 порядков, 27 семейств, около 170 родов и более 1000 видов. Отличаются развитым ценоцитным мицелием непостоянной толщины, в котором септы образуются только для отделения репродуктивных органов.
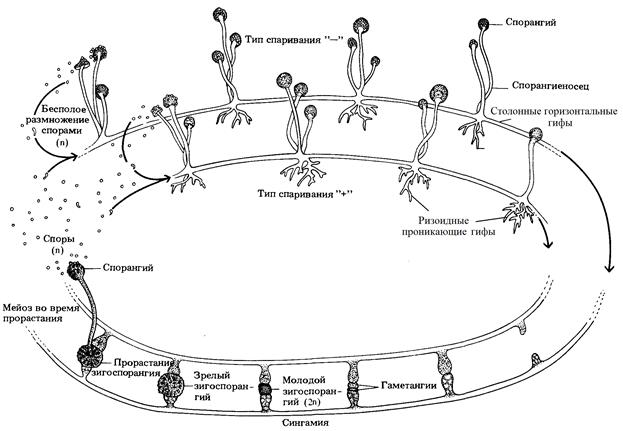
Жизненный цикл развития зигомицетов на примере плесневого гриба мукор
Плесневый гриб мукор прорастает из гетероталличных (разнополых) спор, на субстратах, обогащённых углеводами, образуя одноклеточные гаплоидные многоядерные (без септ) мицелии штаммов «+» и «–» (мицелии грибов не различимы по половым признакам). Вегетативно мукор размножается разрастанием мицелия. Бесполое размножение осуществляется спорами двух типов, конидиями и зигоспорами, которые распространяясь с потоками воздуха на субстрате в благоприятных условиях прорастают в новые мицелии. Конидии – вегетативные споры, образующиеся путём септирования ядер на верхушках выростов мицелия – конидиеносцах. Зигоспоры образуются после осуществления полового процесса. Для этого у зигомицетов, гифы мицелиев штаммов «+» и «–», растут навстречу друг другу. При их контакте, ядра, находящиеся в апикальной (верхушечной) части обоих гиф, изолируются септой от остальной части гифы, образуя клетки – гаметангии. После формирования гаметангии штаммов «+» и «–», сливаются между собой образуя зиготу. Процесс слияния плазмы клеток гаметангиев в зиготе называется плазмогамией, процесс слияния разнополых гаплоидных ядер в диплоидные ядра называется кариогамией и представляет собою половой процесс. Из сформированной зиготы на поверхность субстрата вырастает спорангиеносец. Во время его прорастания диплоидные ядра в спорангиеносце делятся мейозом, образуя генетически рекомбинированные (изменённые) гаплоидные ядра. После выхода на поверхность субстрата на верхушке спорангиеносца образуется зигоспорангий, в котором гаплоидные ядра «+» и «–» изолируются септами в клетках зигоспор. После дозревания зигоспор, зигоспорангий раскрывается и зигоспоры рассыпаются.
Рис. 40. Жизненный цикл развития гриба мукор
Аскомицеты или сумчатые грибы (Ascomycota) – отдел в царстве грибов, объединяющий организмы с септированным (разделённым на части)мицелием и специфическими органами полового спороношения – сумками (асками), содержащими чаще всего по 8 аскоспор. Имеют и бесполое спороношение, причём во многих случаях половой процесс утрачивается, и такие виды грибов относят к несовершенным грибам (Deuteromycota). К аскомицетам относят до 2000 родов и 30 000 видов. Среди них – дрожжи (класс Saccharomycetes) – вторично одноклеточные организмы. Из других известных представителей аскомицетов можно назвать сморчки, пармелию, строчки и трюфели.
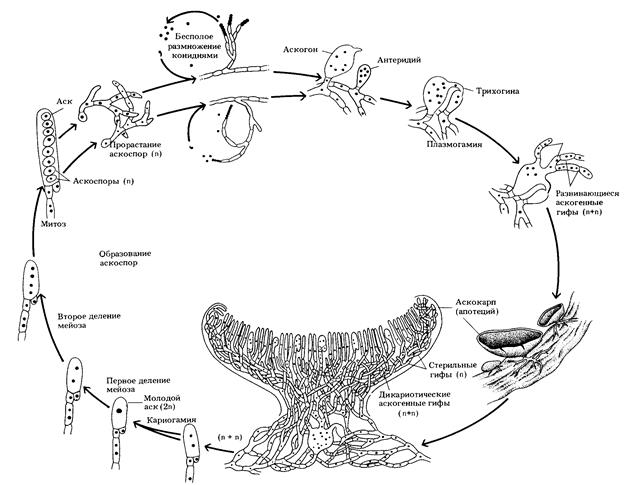
Жизненный цикл развития аскомицетов на примере гриба саркосцифа
Цикл развития грибов аскомицетов начинается с прорастания гетероталличных (разнополых) спор на субстрате в гаплоидные многоклеточные септированные мицелии. Мицелий разрастается в субстрате путём деления апикальных клеток гиф митозом. Для осуществления бесполого размножения спорами, из мицелия гриба вырастают на поверхность субстрата выросты – конидиеносецы на верхушках которых из септированных клеток формируются конидии – вегетативные споры. Для осуществления полового процесса гифы мужского и женского мицелия растут навстречу друг другу. На кончиках этих гиф образуются многоядерные клетки – гаметангии, мужской – антеридий, женский – аскогон. Из аскогона в антеридий растёт трубка – трихогина, по которой ядра из антеридия перемещаются в аскогон. Мужские и женские ядра в аскогоне распределяются парами, но не сливаются между собой. После этого из аскогона начинают расти гифы вторичного мицелия, в которые, по мере их развития, мигрируют пары ядер. В гифах вторичного мицелия и аскогоне проходят синхронные митотические деления, которые приводят к образованию дикарионов – клеток с двумя гаплоидными ядрами (n+n). На поверхности субстрата (у некоторых грибов и внутри субстрата) из первичных гиф мужского и женского мицелиев, а также вторичного дикарионного мицелия формируется плодовое тело – аскокарп. Плодовые тела грибов аскомицетов могут следующих типов: - апотеции – открытые, в виде чаши; - клейстотеции – замкнутые округлённые, из которых аскоспоры выходят после сгнивания или разрушения аскокарпа; - перитеции – колбообразные, с небольшим отверстием для выхода аскоспор. На внутренней поверхности аскокарпа гименофоре, из верхушечных (апикальных) клеток гиф вторичного (аскогенной) дикарионного мицелия – гимения, образуются аски – спорогенные клетки. В клетках асков происходит кариогамия ядер дикариона (n+n), с образованием диплоидного ядра (2n), которое затем делится мейозом и каждое из четырёх гаплоидных ядер ещё раз делится митозом, формируя внутри аска восемь аскоспор (n). У некоторых аскомицетов митотическое деление ядер не происходит, поэтому в аске образуются четыре аскоспоры. После созревания аск лопается и выстреливает споры в воздух, на расстояние 2–30 см. В благоприятных условиях аскоспоры прорастают в субстрат мицелием.
Рис. 41. Жизненный цикл развития гриба аскомицета саркосцифа
Базидиомицеты (Basidiomycota) – отдел царства грибов, включающий виды, производящие споры в булавовидных структурах, именуемых базидии. Вместе с аскомицетами относятся к подцарству высших грибов (Dikarya).
Жизненный цикл развития базидиомицетов на примере шляпочного гриба навозник Цикл развития грибов базидиомицетов начинается с прорастания гетероталличных (разноштаммовых) спор (n) на субстрате в гаплоидные первичные многоклеточные септированные мицелии штамма-плюс «+» и штамма-минус «–». При встрече гиф мицелиев «+» и «–», происходит плазмогамия – срастание двух первичных мицелиев (n) во вторичный дикарионный мицелий (n+n), в каждой клетке которого теперь находится по два ядра «+» и «–». Первичный и вторичный мицелии разрастаются в субстрате путём деления апикальных клеток митозом. Для осуществления бесполого размножения спорами, вторичный мицелий гриба, в местах, обогащённых питательными веществами, формирует плодовое тело – базидиокарп, сформированный из плотно переплетённых гиф мицелия. Само плодовое тело шляпочных грибов состоит из ножки и шляпки. С внутренней стороны шляпки находится гименофор – поверхность, покрытая гимением – слоем спорообразующих клеток. Апикальные спорообразующие клетки базидиомицетов называются базидиями. В базидиях происходит кариогамия – слияние двух «+» и «–» ядер. После кариогамии диплоидное ядро (2n) делится мейозом на четыре гаплоидных ядра (n). На базидии формируются четыре выроста в которые заходят по одному ядру. Выросты с ядрами отделяются от базидии септами (перетяжками), тем самым превращаясь в базидиоспоры (n). После дозревания отделяются от базидии и разносятся воздушными потоками. В благоприятных условиях на субстрате гаплоидные базидиоспоры прорастают в первичные гетероталличные мицелии.
Рис. 42. Жизненный цикл развития гриба базидиомицета
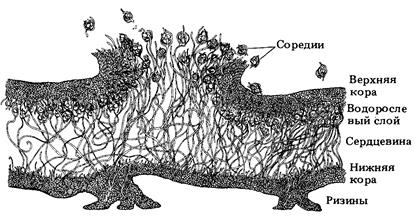
ЛИШАЙНИКИ – симбиотические организмы, состоящие из мицелия грибов аскомицетов и одноклеточных водорослей или цианобактерий. Водоросли или цианобактерии в составе лишайников представлены видами, встречающимися и в свободноживущем состоянии, в то время как лишайниковые грибы, как правило, существуют только в симбиозе с ними. Мицелий гриба поглощает воду с минеральными веществами из окружающей среды и питает ими клетки водорослей. В клетках водорослей происходит фотосинтез, образуются органические вещества, которые поступают в мицелий гриба. Тело лишайника называется слань (рис.). Снаружи слань сформирована плотно переплетёнными желатинизированными гифами мицелия образуя верхнюю и нижнюю кору. Нижняя кора слани тоньше верхней и имеет особые выросты для прикрепления к субстрату – ризины. Внутри сдани находится сердцевина – толстый слой из рыхлых, бесцветных, слабо желатинизированных гиф (этот слой, составляющий 2/3 толщины слани, является запасающим и образован гифами с крупными клетками). Если водоросли в рыхлом мицелии располагаются равномерно, то слань называется гомеомерной, если они расположены одним выраженным водорослевым (гонидиальным) слоем, то – гетеромерной. По форме лишайники бывают: - накипные – плоские, плотно прилегающие к субстрату; - листоватые – прилегающие к субстрату, но имеющие поднимающиеся в виде листовых пластинок края; - кустистые – объёмные, высокие лишайники в виде небольших ветвящихся кустиков. Размножаются лишайники вегетативно изидиями или соредиями – клубочками, в которых гифы мицелия гриба оплетают несколько водорослей. Изидии формируются на поверхности слани как вздутия, после чего отламываются и переносятся водными или воздушными потоками. Соредии формируются внутри слани и вытесняются через поры (отверстия) в коре наружу. Переносимые ветром или водой и попадая в благоприятные условия, изидии или соредии разрастаются в новые лишайники.
Рис. 43. Поперечный срез гетероталомной слани лишайника Lobaria verrucosa
ЖИВОТНЫЕ – царство многоклеточных гетеротрофных организмы способные активно передвигаться. Размножаются вегетативным и половым путём.
РАСТЕНИЯ – царство многоклеточных организмов, преимущественно фотоавтотрофного способа питания. Включает в себя: мхи, папоротники, хвощи, плауны, голосеменные и цветковые растения. Нередко к растениям относят также все водоросли или некоторые их группы. Для растений характерны жизненные формы: деревья, кустарники, полукустарники, травы. Размножаются растения вегетативным (частями вегетативных органов), бесполым (спорами) и половым (семенами) путём.
Мхи или моховидные – отдел высших растений, объеденных в классы: печёночные мхи (6 000–8 000 видов), антоцеротовые мхи (100–200 видов) и листостебельные мхи (10 000 видов). Мхи это, как правило, мелкие растения, длина которых редко превышает 50 мм. Отличаются от других высших растений тем, что в их жизненном цикле взрослые растения представлены разнополыми гаметофитами, а спорофит развивается на женском гаметофите. Размножаются бесполым путём – спорами и вегетативно частями таллома.
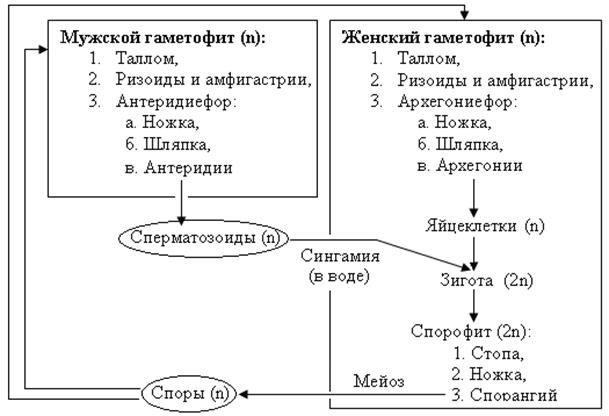
Цикл развития печёночного мха на примере маршанции изменчивой В цикле развития взрослые (многолетние) растения маршанции полиморфной представлены разнополыми (двудомными) мужскими и женскими гаметофитами. Вегетативное тело растения – пластинчатый таллом, имеет вид мясистого лопастного, дихотомически разветвленного слоевища длиной до 10-12 см и шириной до 3 см, образованного множеством клеточных слоев (около 30). Специализированных проводящих тканей таллом не имеет. На нижнем эпидермисе образуются два типа выростов: - ризоиды – бесцветные простые и язычковые одноклеточные выросты, которые выполняют функцию закрепления растения в субстрате. Воду маршанция способна поглощать всем телом, - амфигастрии – окрашенные (темно-фиолетовые) однослойные, многоклеточные чешуи (редуцированные «листья» – филоиды), которые способны долго удерживать воду во время засухи. Различить мужской и женский гаметофиты возможно во время развития на поверхности талломов специальных органов (подставок для гаметангиев) – мужских антеридиефоров и женских архегониефоров. Антеридиефор представляет собой ножку с расположенным сверху восьмиугольным диском (шапкой). С верхней стороны шапки в антеридиальных камерах находятся мужские гаметангии – антеридии. В антеридиях образуются мужские двужгутиковые гаметы – сперматозоиды. Архегониефоры развиваются на женских гаметофитах в виде девятилучевого щитка (звездочки) на ножке. Во время развития архегониефора, архегонии (женские гаметангии) закладываются с морфологически верхней стороны подставки. В силу неравномерного роста обеих сторон ложа подставки (щитка) они смещаются между лучами щитка на его нижнюю сторону, где располагаются группами. Вокруг каждой группы архегониев образуется специальная защитная обертка. Архегонии представляют собой кувшиноподобные камеры, направленные горловиной вниз, в брюшке которых образуется по одной яйцеклетке. Половой процесс в цикле развития маршанции происходит посредством воды преимущественно во время дождя. Капли дождя выбивают из антеридиев зрелые сперматозоиды и вместе с ними отражаются на женские шапки архегониефоров. Сбегая между лучами женских подставок, капли воды вместе со сперматозоидами зависают в месте расположения архегониев, покрывая их. По воде мужские гаметы проникают через шейку архегония в брюшко – калиптру, где оплодотворяют яйцеклетку, образуя зиготу (первую диплоидную клетку спорофита). После оплодотворения вокруг каждого архегония из его ножки начинает образовываться индивидульная чашевидная защитная обертка – 4-5 лопастной псевдоперианций. В это время зигота делится митозом, формируя зародыш спорофита, который прикрепляется к материнскому телу (щитку архегониефора) присоской – гаусторией, и получает оттуда питание. С увеличением размеров зародыша калиптра (брюшко архегония) растягивается (увеличивается в размерах). Взрослый спорофит, развивающийся из зародыша, включает в себя: стопу (присоску) – закрепляющую часть, ножку и спорангий (коробочку спорогона). В процессе своего роста спорофит разрывает калиптру и выносит спорангий наружу. В спорангии формируются диплоидные клетки спорогенной ткани, которые получают питание через удлиненные клетки со спирально утолщенными клеточными стенками – элатеры. В результате разделения спорогенных клеток мейозом образуются гаплоидные гетероталличные споры. При созревании спор элатеры высыхают и скручиваются спирально, разрыхляя массу спор в спорангии. Однослойная коробочка спорангия при созревании спор легко раскрывается восемью загнутыми назад зубцами, а пружинки элатер способствуют равномерному рассеиванию спор. Гетероталличные (разнополые) споры маршанции в благоприятных условиях прорастают в протонемы (передростки) – небольшие беспигментные нитчатые выросты, с верхушечной клетки которых образуются талломы соответственно мужского и женского гаметофита нового поколения. Вегетативное размножение маршанции происходит с помощью выводковых почек, которые формируются в специальных выростах – выводковых корзинках, на верхней стороне как женских, так и мужских талломов. В результате попадания воды на таллом выводковые почки вымываются (распрыскиваются) и прорастают на почве в новые талломы соответствующего пола. Маршанция распространена в увлажненных местах на берегах водоемов, под покровом леса.
Рис. 44. Схема жизненного цикла мха маршанция
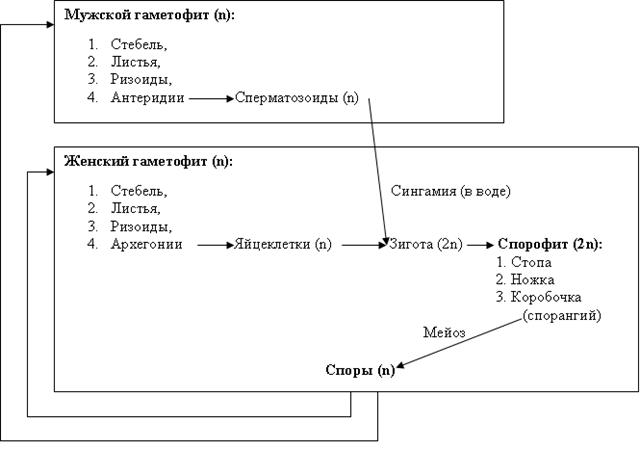
Рис. 45. Жизненный цикл мха маршанция Цикл развития листостебельного мха на примере мха кукушкин лен В цикле развития взрослые растения мха представлены раздельнополыми гаметофитами. Растения кукушкин лен имеют вертикальный стебель, длиной до 40 см, со спирально расположенными в три ряда редуцированными листьями, закрепленный в почве ризоидами. В центре стебля проходит концентрический проводящий тяж, среднюю часть которого занимают водопроводные клетки – гидроиды (восходящий поток), а периферийную – липтоиды – проводящие клетки органических соединений (нисходящий поток). На верхушке мужских растений находятся антеридии, в которых происходит формирование сперматозоидов, на верхушках женских – архегонии с яйцеклетками. Оплодотворение происходит с помощью капель воды. Выбивая из антеридиев сперматозоиды, капли дождя отражаются на архегонии соседних растений. По воде сперматозоиды проникают в брюшко архегониев, где оплодотворяют яйцеклетки. Из оплодотворенной яйцеклетки развивается спорофит, который своей стопой врастает в гаметофит. Ножка спорофита в процессе роста вытягивается в длину на 15-20 см, на верхушке которой образуется коробочка со спорангиями, закрытая крышкой. Край коробочки часто имеет зубчатый край, называемый перистомом. Вызревание спорофита длится до 6 месяцев (у других мхов до 18). В спорангиях спорофита происходит формирование спор из клеток спорогенной ткани, путем деления последних мейозом. После созревания спор крышка коробочки открывается, и споры рассеиваются через перистом. В благоприятных условиях споры прорастают в протонемы – нитевидные хлорофилльные выросты (таллом). Из пазушных почковидных структур протонем вырастают гаметофиты следующего поколения соответствующего пола.
Рис. 46. Схема жизненного цикла мха кукушкин лён
Рис. 47. Жизненный цикл мха кукушкин лён Хвощевидные – отдел высших споровых растений, в настоящее время отнесён в подотдел Хвощёвые отдела Папоротниковидные и насчитывает 15 видов. Хвощёвые характеризуются наличием побегов, состоящих из чётко выраженных метамеров (междоузлий и узлов с мутовчатым расположением листьев). Спорофиты в анатомическом строении напоминают строение покрытосеменных злаковых растений. Бесполое размножение осуществляется разнополыми спорами, вегетативное – корневищами (подземными побегами). Цикл развития хвощёвых на примере хвоща полевого
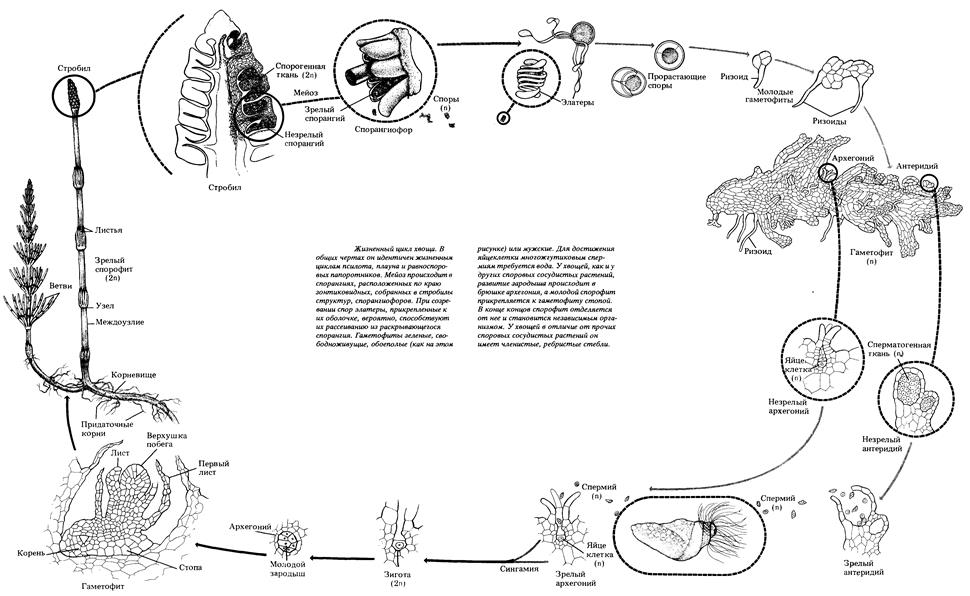
В цикле развития хвоща полевого преобладает спорофит – взрослое, многолетнее растение, состоящее из корневища, закрепленного в почве придаточными корнями. Весной из почек корневища на поверхность почвы вырастает спороносный, бесхлорофилльный вертикальный побег (стебель) с мутовчатым расположением редуцированных (маленьких) листьев, заканчивающийся спороносным колоском (стробилом). Сам спороносный колосок в своем строении имеет ось, на которой размещены зонтиковидные спороносные листья (щитки на ножках) – спорофиллы. С нижней стороны спорофиллов, обращенной к оси стробила, находится от 5 до 10 спорангиев. В спорангиях в результате редукционного деления мейозом клеток спорогенной ткани (2n) образуются гаплоидные, морфологически одинаковые, но разнополые споры (мужские и обоеполые). Оболочки спор имеют специальные выросты – элатеры, которые скручены спирально вокруг спор во влажном состоянии, а при подсыхании разворачиваются. Это позволяет спорам цепляться друг за друга и распространяться группами. После созревания спор спороносные листья стробила размыкаются, спорангии лопаются и споры разносятся ветром. Вследствие содержания в спорах хлоропластов они быстро (в течение 3-х недель) теряют всхожесть. Попадая на увлажненный илистый грунт, группы спор прорастают в хлорофиллоносные гаметофиты в виде лопастных пластинок, закрепляющихся в субстрате ризоидами. Гаметофиты достигают половой зрелости через 3-5 недель после прорастания. На мужских гаметофитах, меньшего размера, образуются антеридии – мужские гаметангии, в которых образуются многожгутиковые сперматозоиды. На двуполых гаметофитах, более рассеченной формы, архегонии (женские гаметангии) развиваются раньше антеридиев, что увеличивает вероятность перекрестного оплодотворения. Для достижения сперматозоидами яйцеклеток, находящихся в архегониях, необходима вода. На одном гаметофите могут быть оплодотворены сразу несколько яйцеклеток, из которых в дальнейшем развиваются зародыши, есть молодые спорофиты. Зародыши крепятся стопой к животику архегония и получают из гаметофита необходимые питательные вещества для развития, формируя зачаточный корешок, стебель и почку. После формирования, зародышевый корешок начинает расти, закрепляется в почве и молодой спорофит отсоединяется от гаметофита, который через некоторое время отмирает. После спороношения весенние (спороносные) побеги отмирают и из почек корневища вырастают зеленые ассимиляционные побеги. Ассимиляционные побеги имеют вертикальный стебель с мутовчатым расположением на нем боковых ветвей и листьев под ними. Они выполняют функцию образования органических соединений в процессе фотосинтеза и накопления их в корневище. В конце вегетационного периода ассимиляционные побеги отмирают, остается корневище, которое зимует в почве.
Рис. 48. Схема жизненного цикла хвоща полевого
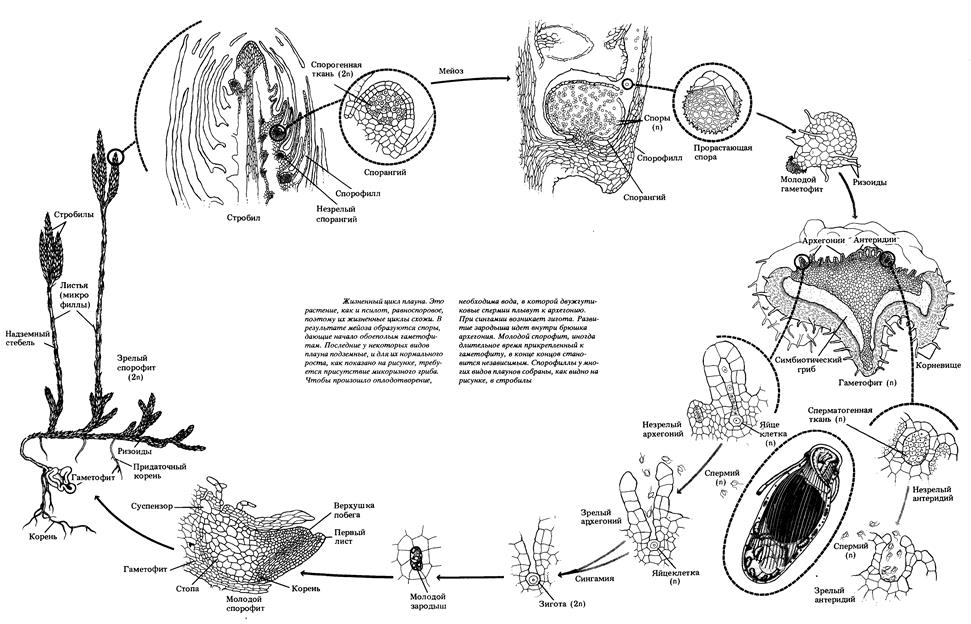
Рис. 49. Жизненный цикл хвоща полевого Плауновидные – отдел высших споровых растений, представлен 1200 видами. Характеризуются наличием развитой проводящей системы, корнепобеговым типом строения спорофитов и талломным строением гаметофитов, бесполым размножением спорами и вегетативным размножением надземными побегами. Цикл развития плауновидных растений на примере плауна булавовидного Спорофит плауна булавовидного – многолетнее вечнозеленое растение, преобладает в цикле развития. В своем составе имеет стелющийся дихотомически разветвленный стебель, покрытый спирально расположенными маленькими ланцетно-линейными листочками (микрофиллами) и закрепленный в почве дихотомически разветвленными корнями. Стеблевые побеги заканчиваются верхушечными почками или спороносными колосками (стробилами). Спороносные колоски на длинных ножках состоят из оси, на которой спирально расположены спороносные листья (спорофиллы) со спорангиями с верхней (дорсальной) стороны. После редукционного (мейотического) деления материнских диплоидных клеток образуются гаплоидные споры. Споры покрыты двойной оболочкой (эндо- и экзоспорием) и содержат до 50% масла. После созревания споры рассеиваются и в благоприятных условиях прорастают. Развитие гаметофита (заростка) из спор происходит медленно в течение 12-20 лет. Он образует ризоиды, которыми закрепляется в почве и поглощает воду, а также вступает в симбиоз с мицелием гриба, который размещается в его коровой части. Заросток развивается в почве без доступа света, поэтому не имеет хлоропластов, однако если он выходит на поверхность, они образуются. Гаметофит двуполый и напоминает луковицу, позже разрастается до 2-3 см в диаметре, и имеет блюдцеобразную форму. Антеридии и архегонии размещены рядом на верхней стороне и погружены в паренхимную ткань. Антеридии овальной формы, архегонии – колбообразные. В брюшной части архегониев содержится яйцеклетка и брюшная канальцевая клетка, в шейке – шейные канальцевые клетки. Для перемещения сперматозоидов с антеридиев к яйцеклеткам архегониев необходима вода. Заплывая в шейку архегония, сперматозоид движется в брюшко архегония, где сливается с яйцеклеткой, образуя зиготу – первую диплоидную клетку спорофита. Зародыш – молодой спорофит, медленно развивается на гаметофите, получая из него необходимые питательные вещества. Когда корни зародыша прорастают в почву, он отсоединяется от гаметофита и разрастается во взрослое растение. При постепенном созревании архегониев на гаметофите в них могут образовываться и развиваться несколько разновозрастных спорофитов одновременно.
Рис. 50. Схема жизненного цикла плауна булавовидного
Рис. 51. Жизненный цикл плауна булавовидного Папоротникообразные – отдел высших споровых растений, объединяющий около 11 000 видов. Папоротникообразные растения характеризуются корнепобеговым строением тела спорофитов и талломным строением тела гаметофитов. Размножаются бесполым путём – спорами и вегетативно – побегами (корневищами). Популярное:
|
Последнее изменение этой страницы: 2016-06-04; Просмотров: 8153; Нарушение авторского права страницы