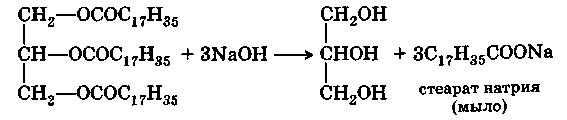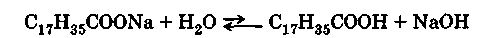|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Очистка белков избирательной денатурациейСтр 1 из 7Следующая ⇒
Методы очистки белков Наиболее трудоёмкий этап получения индивидуальных белков - их очистка от других белков, находящихся в растворе, полученном из данной ткани. Часто изучаемый белок присутствует в небольших количествах, составляющих доли процента от всех белков раствора. Так как белки обладают конформационной лабильностью, при работе с белками следует избегать денатурирующих воздействий, поэтому выделение и очистка белков происходят при низких температурах. На первых стадиях очистки белков целесообразно использовать методы, учитывающие какую-либо характерную особенность данного белка, например термостабильность или устойчивость в кислых растворах. Первыми методами очистки необходимо удалить из раствора основную массу балластных белков, которые значительно отличаются от выделяемого белка физико-химическими свойствами. Впоследствии применяют всё более тонкие методы очистки белка. Очистка белков избирательной денатурацией Большинство белков денатурирует и выпадает в осадок уже при кратковременном нагревании раствора до 50-70°С или подкислении раствора до рН 5. Если выделяемый белок выдерживает эти условия, то с помощью избирательной денатурации можно удалить большую часть посторонних белков, отфильтровав выпавшие в осадок-белки, или осадить их центрифугированием. Высаливание Метод очистки белков, основанный на различиях в их растворимости при разной концентрации соли в растворе. Соли щелочных и щёлочно-земельных металлов вызывают обратимое осаждение белков, т.е. после их удаления белки вновь приобретают способность растворяться, сохраняя при этом свои нативные свойства. Чаще всего для разделения белков методом высаливания используют разные концентрации солей сульфата аммония - (NH4 )2 SO4 . Чем выше растворимость белка, тем большая концентрация соли необходима для его высаливания. Гель-фильтрация, или метод молекулярных сит Для разделения белков часто используют хроматографические методы, основанные на распределении веществ между двумя фазами, одна из которых подвижная, а другая неподвижная. В основу хроматографических методов положены разные принципы: гель-фильтрации, ионного обмена, адсорбции, биологического сродства. Метод разделения белков с помощью гель-фильтрационной хроматографии основан на том, что вещества, отличающиеся молекулярной массой, по-разному распределяются между неподвижной и подвижной фазами. Хроматографическая колонка заполняется гранулами пористого вещества (сефадекс, агароза и др.). В структуре полисахарида образуются поперечные связи и формируются гранулы с "порами", через которые легко проходят вода и низкомолекулярные вещества. В зависимости от условий можно формировать гранулы с разной величиной "пор". Неподвижная фаза - жидкость внутри гранул, в которую способны проникать низкомолекулярные вещества и белки с небольшой молекулярной массой. Смесь белков, нанесённую на хроматографическую колонку, вымывают (элюируют), пропуская через колонку растворитель. Вместе с фронтом растворителя движутся и самые крупные молекулы. Более мелкие молекулы диффундируют внутрь гранул сефадекса и на некоторое время попадают в неподвижную фазу, в результате чего их движение задерживается. Величина пор определяет размер молекул, способных проникать внутрь гранул . Так как гелевая структура сефадекса легко деформируется под давлением, гели стали заменять более жёсткими матрицами (сефактил, той-оперл), представляющими сферические гранулы с разными размерами пор. Выбор размеров пор в гранулах зависит от целей хроматографии (о других хроматографических методах будет сказано ниже). Ультрацентрифугирование Метод разделения также основан на различии в молекулярных массах белков. Скорость седиментации веществ в процессе вращения в ультрацентрифуге, где центробежное ускорение достигает 100 000-500 000 g, пропорционально их молекулярной массе. На поверхность буферного раствора, помещённого в кювету, наносят тонкий слой смеси белков. Кювету помещают в ротор ультрацентрифуги. При вращении ротора в течение 10-12 ч более крупные молекулы (с большей молекулярной массой) оседают в буферном растворе с большей скоростью. В результате в кювете происходит расслоение смеси белков на отдельные фракции с разной молекулярной массой .После расслоения белковых фракций дно кюветы прокаливают иглой и по каплям собирают содержимое небольшими порциями в пробирки.
Электрофорез белков Метод основан на том, что при определённом значении рН и ионной силы раствора белки двигаются в электрическом поле со скоростью, пропорциональной их суммарному заряду. Белки, имеющие суммарный отрицательный заряд, двигаются к аноду (+), а положительно заряженные белки - к катоду (-). Электрофорез проводят на различных носителях: бумаге, крахмальном геле, полиакриламидном геле и др. В отличие от электрофореза на бумаге, где скорость движения белков пропорциональна только их суммарному заряду, в полиакриламидном геле скорость движения белков пропорциональна их молекулярным массам. Разрешающая способность электрофореза в полиакриламидном геле выше, чем на бумаге. Так, при электрофорезе белков сыворотки крови человека на бумаге обнаруживают только 5 главных фракций: альбумины, α1 глобулины, α2 -глобулины, β-глобулины и γ-глобулины (рис. 1-57). Электрофорез тех же белков в полиакриламидном геле позволяет получить до 18 различных фракций. Для обнаружения белковых фракций полоски бумаги или столбики геля обрабатывают красителем (чаще всего бромфеноловым синим или амидовым чёрным). Окрашенный комплекс белков с красителем выявляет расположение различных фракций на носителе.
2. Назовите три-четыре сходства ферментов c небиологическими катализаторами. Ферменты и небиологические катализаторы, подчиняясь общим законам катализа, имеют следующие сходные признаки. 1. Они катализируют только энергетически возможные реакции. 2. Они никогда не изменяют направления реакции. 3. Они не изменяют равновесия обратимой реакции, а лишь ускоряют его наступление. 4. Они не расходуются в процессе реакции. Поэтому фермент в клетке работает до тех пор, пока по каким - либо причинам не разрушится. Однако ферменты обладают и особыми качествами, отличающими их от небиологических катализаторов. Эти отличия связаны с особенностями строения ферментов, являющихся сложными белковыми молекулами. 1. Скорость ферментативного катализа намного выше, чем небиологического. Из этого следует, что ферменты сильнее снижают энергию активации реакции, чем небиологические катализаторы. 2. Ферменты обладают высокой специфичностью. Есть ферменты, действующие только на один из стереоизомеров вещества, тогда как платина, например, используется в качестве катализатора при самых разнообразных реакциях. Высокая специфичность позволяет ферментам направлять обмен веществ в строгое русло. 3. Ферменты катализируют химические реакции в « мягких» условиях, т.е. при обычном давлении, невысокой температуре ( около 37оС) и рН среды, близком к нейтральной. Это отличает их от других катализаторов, действующих при больших давлениях, крайних значениях рН и высокой температуре. Ферменты из-за белкового строения весьма чувствительны к изменениям температуры, т.е. термолабильны, и к сдвигам рН среды. 4. Ферменты являются катализаторами с регулируемой активностью, чего нельзя сказать о небиологических катализаторах. Это уникальное свойство ферментов позволяет изменять скорость превращения веществ в организме в зависимости от условий среды, т.е. приспосабливаться к действию различных факторов. 5. Скорость ферментативной реакции прямо пропорциональна количеству фермента, тогда как для небиологического катализа не существует строгой зависимости скорости реакции от количества катализатора. Поэтому недостаток фермента в живом организме означает низкую скорость превращения вещества и , наоборот, одним из путей приспособления клеток организма является образование дополнительных количеств фермента.
3. Приведите три-четыре примера коферментов. ТИАМИНОВЫЕ в составе витамин В1 (ТИАМИН) - ТМФ – ТИАМИНМОНОФОСФАТ, ТДФ- ТИАМИНДИФОСФАТ, ТТФ - ТИАМИНТРИФОСФАТ. ТПФ связана с ферментами ДЕКАРБОКСИЛАЗАМИ альфа КЕТОКИСЛОТ (ПВК, альфа КГК)
2.ФЛАВИНОВЫЕ содержат витамин В2 - ФМН – ФЛАВИНМОНОНУКЛЕОТИД, ФАД - ФЛАВИИАДЕНИНДИНУКЛЕОТИД. ФМН и ФАД связанны с ферментами ДЕГИДРОГЕНАЗАМИ. Участвуют в реакциях ДЕГИДРИРОВАНИЯ. 3. ПАНТОТЕИНОВЫЕ (витамин ВЗ) - KOF A (HS-KOA - HS КОЭНЗИМ А) - КОФЕРМЕНТ АЦИЛИРОВАНИЯ. 4. НИКОТИНАМИДНЫЕ содержат витамин РР (НИАЦИН)- НАД (НИКОТИНАМИДАДЕНИНДИНУКЛЕОТИД), НАДФ (НИКОТИНАМИДАДЕНИНДИНУКЛЕОТИДФОСФАТ). Связаны с ДЕГИДРОГЕНАЗАМИ:
4. Назовите три основных проявления дефицита витамина PP (три «Д»). пожилой возраст; регулярное соблюдение строгого диетического питания; чрезмерное употребление спиртных напитков и наркотических веществ. 6. Назовите основную функцию ДНК полимеразы. Задача ДНК-полимеразов с биологической точки зрения заключена в синтезе ДНК и касается его катализа. Фактически общие ДНК полимераза параметры данных ферментов одинаковые. Но все же конкретный образец ДНК полимераза характерный своими отличительными свойствами. Последние завязаны на определенной биологической функции. Она должна реализовываться благодаря структурным строениям. Ученые выделяют весьма немалую цифру ДНК-полимераз. Касательно человеческих клеток удалось идентифицировать репликативные полимеразы, ДНК-полимераза бета и гамма. ДНК полимераза и её идентификация помогла открыть в некоторых специфических человеческих клетках так называемые транскриптазы, которые впоследствии назвали обратными. На сегодня роль днк полимераза остается неизвестной. Кроме того удалось значительно скрупулезнее изучить и иной фермент – дезоксинуклеотидилтрансферазу. Ее функции, по убеждению исследователей, завязаны на синтезе антител. 7. Напишите реакцию катализируемую каталазой. Каталаза, гемсодержащий фермент, присутствует в пероксисомах всех клеток человека и обладает чрезвычайно высокой молекулярной активностью. В эритроцитах она находится в цитозоле и защищает гемоглобин от окисления. РАСПАД ГЛИКОГЕНА Распад гликогена (гликогенолиз) осуществляется в периоды между приёмами пищи. В это время печень расщепляет находящийся в ней гликоген с определенной скоростью, которая позволяет организму сохранять концентрацию глюкозы в крови на неизменном уровне. СИНТЕЗ И РАСПАД ГЛИКОГЕНА Синтез и распад гликогена регулируются посредством нервной системы и гормонов. Это - два самостоятельных процесса, проходящих различными путями. Как мы уже рассмотрели, основная роль гликогена - регулирование концентрации глюкозы в крови, а также создание того резерва глюкозы, который необходим для интенсивной мышечной работы.
5. Аэробный распад глюкозы. Биологическая роль, схема, конечные продукты, ключевые ферменты. 6. Гормональная регуляция уровня глюкозы крови. Эффекты инсулина, глюкагона, кортизола и адреналина на процессы обмена глюкозы в тканях-мишенях. Нарушения углеводного обмена при сахарном диабете. Гор регул. уровня глюк. в крови. Гипер и гипогликемические гормоны. Глюкагон, кортизол, адреналин, хим природа и место синтеза, тк-мишени, эффекты на углеводный обмен в тк-мишенях. · Глюкагон связывается с рецептором на плазматической мембране и активируется при посредничестве G -белка аденилатциклазу, катализирует образование АМФ и АТФ. Далее в печени проис. активация гликогенфосфорилазы и ингибировние гликогенсинтетезы. Это приводит к высвобождению из гликогена глюкозо-1-фосфата, который прев. в глюкозо-6 -фосфат, под влиянием глюкозо-6 фосфотазы образуется свободная глюкоза, способная выйти из клетки в кровь. · Адреналин: Стимулирует выделение глюкозы из печени в кровь чтобы снабдить тк. "топливом" в экстрим. ситуации. Эффект адреналина в печени обусловлен фосфорилированием гликогенфосфорилазы. · Система передачи сигнала в кл. зависит от типа рецептора, с которым взаимодейст адреналин. · Пример: Взаимодействие адрен. с бетта 2 -рецептором кл. печени приводит в действие аденилатциклазную систему. А с альфа1 -рец включает инозитолфосфатный механизм трансмемб. передачи гормонального сигнала. · Результат действия обоих систем фосфорилирование ключевых ферментов и переключ. прод. синтеза гликогена и его распад. · Кортизол - синтезируется из холестерола, который поступает из крови в составе ЛПНП или синтезируется в клет.из АЦЕТИЛ-КоА. · Скорость синтеза и секреции кортизола стимулируется в ответ на стресс "травму" повышение к-ции кортизола подавляет синтез кортиколиберина и АКТГ по мех. отриц. обратной связи. · Гипер и гипоглик. Инсулин..... · Гипогликемия - сопровождается уменьшением содерж. инсулина и снижением отношения инсулин -глюкагон. что приводит к ускорению липолиза жировой ткани. · Гормоны ЖКТ - гастрин, секретин, энтероглюкагон. · Гипергликемия -повыш. содер. глюкозы в крови. · Инсулин - это белковый гормон, 51а аминокислота сост из 2х полипептидных ципей. · - Снижается в бетта кл. поджелудочной железы и секретируется в кровь в ответ на повышение конц. глюкозы в крови. · Инсулин стимулирует перемещение ГЛЮТ -4 и встраивание их в мембрану кл. Влияние инсулина на скорость синтеза гликогена в тк. осущ посредств. измен. активности гликогенсинтетазы и гликогенфосфорилазы - ключевых ферментов.
7. Динамическое состояние белков в организме. Азотистый баланс, его виды. Заменимые и незаменимые аминокислоты. Отрицательный азотистый баланс: основные причины развития, метаболические последствия и возможные клинические проявления. Динамическое состояние: Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и внешней средой. Аминокислоты непосредственно участвуют в биосинтезе большого количества других биологически активных соединений, регулирующих процессы обмена веществ в организме, таких как нейромедиаторы и гормоны. Аминокислоты служат донорами азота при синтезе всех азотсодержащих небелковых соединений, в том числе нуклеотидов, гема, креатина, холина и др. 8.Аммиак: пути образования, механизм токсического действия, пути обезвреживания. Орнитиновый цикл мочевинообразования: биологическое значение и химическая сущность процесса, источники атомов мочевины, органная и внутриклеточная локализация ферментов цикла. Взаимосвязь орнитинового и цитратного циклов Кребса. Аммиак постоянно образуется во всех органах и тканях организма. Наиболее активными его продуцентами в кровь являются органы с высоким обменом аминокислот и биогенных аминов – нервная ткань, печень, кишечник, мышцы. Основными источниками аммиака являются следующие реакции: · внутримолекулярное дезаминирование гистидина, катаболизм некоторых аминокислот (серина, треонина, глицина) – в печени, · окислительное дезаминирование глутаминовой кислоты во всех тканях (кроме мышечной), особенно в печени и почках, · дезаминирование амидов глутаминовой и аспарагиновой кислот – в печени и почках, · катаболизм биогенных аминов – во всех тканях, в наибольшей степени в нервной ткани, · жизнедеятельность бактерий толстого кишечника, · распад пуриновых и пиримидиновых оснований – во всех тканях.
9. Реакции декарбоксилирования. Структура декарбоксилаз. Биогенные амины: образование, функции, пути инактивации (на примере гамма-аминомасляной кислоты). Роль гидроксилирования, декарбоксилирования и метилирования в образование катехоламинов. 10. Гормональная регуляция белкового обмена. Эффекты соматотропного гормона, половых гормонов, глюкокортикоидов, инсулина.. Приступая к изложению материала по белковому метаболизму, уместно напомнить его гормональную регуляцию. Основными гормонами, регулирующими белковый обмен, являются СТГ, половые стероиды, тиреоидные (Т3, Т4), пептидные гормоны островков Лангерганса – глюкагон и инсулин, а также глюкокортикоиды и нейропептид лептин. Соматотропин (СТГ) способствует синтезу белка в висцеральном (внутренние органы) и соматическом отсеках (скелетные мышцы, кости, хрящи), а также в лимфоидных органах. Механизм действия СТГ состоит в ускорении утилизации аминокислот клетками. Соответственно, при акромегалии и гипофизарном гигантизме наблюдается положительный азотистый баланс, при гипофизэктомии и гипофизарном нанизме – отрицательный. СТГ относится к контринсулярным гормонам. Под его влиянием повышается продукция инсулиновых рецепторов и высвобождается инсулин и глюкагон из островков Лангерганса. В то же время, на белковый обмен СТГ действуют как синергист инсулина, обеспечивая анаболический эффект. СТГ использует для этой цели энергию катаболизма жиров, а инсулин – за счет окисления экзогенной глюкозы. Дефицит инсулина тормозит анаболический эффект СТГ. СТГ способствует анаболическому действию андрогенов. В свою очередь, тиреоидные гормоны, инсулин, андрогены пермиссивно способствуют эффектам СТГ на метаболизм протеинов. Инсулин. Механизмы его анаболического действия следующие: • стимулирует сборку рибосом и трансляцию; • усиливает активный транспорт аминокислот в клетки; • тормозит глюконеогенез из аминокислот; • тормозит освобождение аминокислот из мышечных клеток; • усиливает синтез ДНК, РНК и митотическую активность инсулинозависимых тканей. При инсулинозависимом сахарном диабете (ИЗСД) угнетается синтез белка, усиливается его катаболизм, особенно в скелетных мышцах, формируются гипераминоацидемия, отрицательный азотистый баланс, повышается выделение мочевины с мочой. Эти факторы способствуют возникновению вторичного иммунодефицита, симптомов мышечной слабости. Выработка инсулина активируется тиреоидными гормонами, тормозится глюкокортикоидами, хотя и те, и другие являются антагонистами его эффектов. Секреция инсулина стимулируется белковой пищей, аргинином и лейцином. Глюкагон. Он тормозит поглощение аминокислот, синтез белка, усиливает протеолиз и освобождение аминокислот мышцами. В печени он стимулирует глюконеогенез из аминокислот и кетогенез. Глюкагон является антагонистом анаболических эффектов инсулина и СТГ. Продукция его в островках Лангерганса стимулируется аргинином и аланином, тиреоидными гормонами, глюкокортикоидами, СТГ. Гиперглюкагонемия наблюдается при ИЗСД, опухолевом росте – глюкагономе, голодании. Соматостатин. Он тормозит синтез белка без усиления его распада, является антагонистом инсулина, глюкагона, СТГ. Его высвобождение стимулирует выработку СТГ, тироксин и глюкокортикоиды. Глюкокортикоиды. Тормозят синтез белка и увеличивают путем глюконеогенеза его распад в лимфоидной ткани и соматическом отсеке, но не в печени, где биосинтез многих глобулинов и трансаминаз, наоборот, усиливается. В этом качестве они участвуют в переброске аминокислот из соматического отсека в висцеральный во время стресса, голодания, ответа острой фазы (ООФ), или синдрома системной воспалительной реакции (ССВР), травматических повреждений. Однако синтез печеночных белков не уравновешивает их распад в соматическом отсеке, поэтому при гиперкортицизме (болезнь и синдром Иценко-Кушинга) имеются отрицательный азотистый баланс, гипераминоацидемия, аминоацидурия. Этот механизм обусловливает клинический симптомокомплекс – атрофию мышц, истончение кожи, гипоплазию тимико-лимфатического аппарата, остеопороз. При раннем дебюте заболевания тормозится рост больных. Действие глюкокортикоидов антагонистическое по отношению к СТГ и инсулину, синергичное – к глюкагону. 11. Липиды. Строение, классификация. Общие закономерности структуры и функции. ЛИПИДЫ - это разнородная группа природных соединений, полностью или почти полностью нерастворимых в воде, но растворимых в органических растворителях и друг в друге, дающих при гидролизе высокомолекулярные жирные кислоты. В живом организме липиды выполняют разнообразные функции. Биологические функции липидов: Структурная Структурные липиды образуют сложные комплексы с белками и углеводами, из которых построены мембраны клетки и клеточных структур, участвуют в разнообразных процессах, протекающих в клетке. Запасная (энергетическая) Запасные липиды (в основном жиры) являются энергетическим резервом организма и участвуют в обменных процессах. В растениях они накапливаются главным образом в плодах и семенах, у животных и рыб — в подкожных жировых тканях и тканях, окружающих внутренние органы, а также печени, мозговой и нервной тканях. Содержание их зависит от многих факторов (вида, возраста, питания и т. д.) и в отдельных случаях составляет 95—97% всех выделяемых липидов. Калорийность углеводов и белков: ~ 4 ккал/грамм. Калорийность жира: ~ 9 ккал/грамм. Преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность – он не связан с водой. Это обеспечивает компактность жировых запасов - они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме – примерно 400 гр.; при голодании этого количества не хватает даже на одни сутки. Защитная Подкожные жировые ткани предохраняют животных от охлаждения, а внутренние органы — от механических повреждений. Образование запасов жира в организме человека и некоторых животных рассматривается как приспособление к нерегулярному питанию и к обитанию в холодной среде. Особенно большой запас жира у животных, впадающих в длительную спячку (медведи, сурки) и приспособленных к обитанию в условиях холода (моржи, тюлени). У плода жир практически отсутствует, и появляется только перед рождением. Особую группу по своим функциям в живом организме составляют защитные липиды растений — воски и их производные, покрывающие поверхность листьев, семян и плодов. Классификация липидов Общепринятой классификации липидов не существует. Наиболее целесообразно классифицировать липиды в зависимости от их химической природы, биологических функций, а также по отношению к некоторым реагентам, например, к щелочам. По химическому составу липиды обычно делят на две группы: простые и сложные. Простые липиды – сложные эфиры жирных кислот и спиртов. К ним относятся жиры, воски и стероиды. Жиры – эфиры глицерина и высших жирных кислот. Воски – эфиры высших спиртов алифатического ряда (с длинной углеводной цепью 16-30 атомов С) и высших жирных кислот. Стероиды – эфиры полициклических спиртов и высших жирных кислот. Сложные липиды – помимо жирных кислот и спиртов содержат другие компоненты различной химической природы. К ним относятся фосфолипиды и гликолипиды. Фосфолипиды – это сложные липиды, в которых одна из спиртовых групп связана не с ЖК, а с фосфорной кислотой (фосфорная кислота может быть соединена с дополнительным соединением). В зависимости от того, какой спирт входит в состав фосфолипидов, они подразделяются на глицерофосфолипиды (содержат спирт глицерин) и сфингофосфолипиды (содержат спирт сфингозин). Гликолипиды – это сложные липиды, в которых одна из спиртовых групп связана не с ЖК, а с углеводным компонентом. В зависимости от того, какой углеводный компонент входит в состав гликолипидов, они подразделяются на цереброзиды (в качестве углеводного компонента содержат какой-либо моносахарид, дисахарид или небольшой нейтральный гомоолигосахарид) и ганглиозиды (в качестве углеводного компонента содержат кислый гетероолигосахарид). Иногда в самостоятельную группу липидов (минорные липиды) выделяют жирорастворимые пигменты, стерины, жирорастворимые витамины. Некоторые из этих соединений могут быть отнесены к группе простых (нейтральных) липидов, другие — сложных. По другой классификации липиды в зависимости от их отношения к щелочам делят на две большие группы: омыляемые и неомыляемые. К группе омыляемых липидов относятся простые и сложные липиды, которые при взаимодействии со щелочами гидролизуются с образованием солей высокомолекулярных кислот, получивших название «мыла». К группе неомыляемых липидов относятся соединения, не подвергающиеся щелочному гидролизу (стерины, жирорастворимые витамины, простые эфиры и т. д.). По своим функциям в живом организме липиды делятся на структурные, запасные и защитные. Структурные липиды - главным образом фосфолипиды. Запасные липиды - в основном жиры. Защитные липиды растений — воски и их производные, покрывающие поверхность листьев, семян и плодов, животных – жиры.
12. Гликолипиды (цереброзиды, ганглиозиды). Холестерин: строение, синтез. Биологическая роль. Гликолипиды - сложные липиды, одна из спиртовых групп которых связана с углеводным компонентом. Состоят из сфингозина, жирной кислоты и молекулы какого-либо углевода. Если в формулу СФЛ вместо фосфорной кислоты поставить какой-нибудь углевод, то получим формулу ГЛ. Гликолипиды тоже имеют гидрофильную "головку" и 2 гидрофобных "хвоста". Общая схема их строения представлена на рисунке:
Чаще всего в построении молекулы гликолипидов участвуют D-галактоза, D-глюкоза, уроновые кислоты:
1. ЦЕРЕБРОЗИДЫ. В качестве углеводного компонента содержат какой-либо моносахарид (глюкоза, галактоза), либо дисахарид, или нейтральный небольшой олигосахарид. 2. ГАНГЛИОЗИДЫ. Углеводным компонентом является олигосахарид, состоящий из разных мономеров, как самих моносахаридов, так и их производных. Этот олигосахарид обязательно кислый, в его состав обязательно входит сиаловая кислота. Благодаря определенной последовательности мономеров, олигосахариды в составе ганглиозида придают молекуле выраженные антигенные свойства
Омыление жиров с помощью гидроксида натрия или гидроксида калия проводится главным образом при получении мыла: Мыла представляют собой щелочные соли высших жирных кислот:
В промышленности в качестве исходных веществ для и получения применяются животные жиры (сало низких сортов), хлопковое, пальмовое, кокосовое масла, гидрогенизированные жиры. При нагревании их с гидроксидом натрия образуется густой раствор («мыльный клей»), содержащий глицерин и соли жирных кислот. Затем к еще горячей жидкости прибавляют поваренную соль — «высаливают» натриевое мыло. Натриевые мыла после застывания представляют собой твердую массу. Мягкие, или жидкие, мыла являются обычно калиевыми мылами. Их получают из менее ценных жиров (льняного, конопляного масла, ворвани) путем омыления гидроксидом калия, но при этом не производят технически слишком дорогого выделения калиевых солей жирных кислот, в результате чего калиевые мыла содержат еще воду и глицерин. Все мыла, являясь щелочными солями слабых кислот, в воде частично гидролизуются с образованием свободной жирной кислоты и гидроксида щелочного металла, поэтому их растворы имеют щелочную реакцию:
При добавлении натриевого мыла к жесткой воде ионы кальция и магния замещают ионы натрия, образуя нерастворимые и поэтому не обладающие моющим действием кальциевые и магниевые мыла. Поэтому моющая способность мыла в жесткой воде значительно снижается. В продаже под различными названиями имеются моющие средства (детергенты), заменители мыл, представляющие собой, например, смесь натриевых солей эфиров серной кислоты и высших спиртов. Эти соединения можно использовать и в жесткой воде, так как они не образуют нерастворимых соединений с кальцием и магнием; они не обладают сильными щелочными свойствами и поэтому не повреждают тканей. Мыла и детергенты представляют собой эмульгирующие вещества, превращающие смесь масла и воды в устойчивую эмульсию. Благодаря эмульгированию находящиеся на коже или на одежде жиры и масла, впитывающие в себя грязь, можно удалить водой. Углеводородная часть молекулы мыла, или «хвост», имеет тенденцию растворяться в капле масла, в то время как карбоксильная группа, или «голова», притягивается к водной фазе. Вследствие этого поверхность каждой капли масла приобретает отрицательный заряд и стремится оттолкнуться от других таких же капель масла, что приводит к образованию устойчивой эмульсии.
13. Жирные кислоты, понятие о биосинтезе, заменимые и незаменимые жирные кислоты, их биологическая роль. Незаменимые жирные кислоты (НЖК) называются так, потому что они не могут быть синтезированы в организме человека, и нам нужно потреблять их с пищей. Они играют ключевую роль в здоровье нашей кожи, в росте и развитии, стабильности работы сердца, в свертываемости и течении нашей крови. Избыток, недостаток или неправильное соотношение этих жизненно необходимых веществ может нанести ущерб нашему здоровью. В настоящее время незаменимыми считаются две жирные кислоты: альфа-линоленовая кислота (АЛК) и линолевая кислота (ЛК), омега–3 и омега–6 соответственно. Омега–3 и омега–6 обычно воспринимаются как синонимы НЖК, на самом деле из АЛК и ЛК формируются 12 жирных кислот. И все жирные кислоты, которые происходят от них, классифицированы как омега–3 и омега–6. Таким образом, не все кислоты из семейств омега 3 и 6 являются незаменимыми. Только полиненасыщенные жиры АЛК и ЛК должны поступать из внешних источников. Жирные кислоты, синтезируемые из АЛК, включают в себя такие известные вещества, как эйкозопентаэоновая(ЭПК) и докозагексаэоновая(ДГК) кислоты. В семействе омега–6 самые известные — арахидоновая и гамма-линоленовая кислоты. Интересно, что современные исследования подвергает сомнению незаменимость АЛК, так как найдено подтверждение того, что тело, возможно, в состоянии ее синтезировать самостоятельно. К счастью, нам не стоит беспокоиться об этом, потому что и линолевая и альфа-линоленовая жирные кислоты очень распространены в составе растительных жиров. Никаких официальных рекомендаций по объему потребления НЖК не существует. Но зато нам известно, что их соотношение не менее, а то и более важно, чем их потребляемый объем. Ученые, как правило, сходятся в том, что древний человек потреблял жирные кислоты омега–6 и омега–3 в примерном соотношении 1:1. Именно в таком соотношении незаменимые жирные кислоты присутствуют в мозге человека. Исторически, с ростом потребления зерновых и масел с высоким содержанием омега–6, которые мы часто употребляем в пищу, это соотношение начало изменяться. Современные диетологи считают, что идеальное отношение омега–6 и омега–3 — от 1:1 до 4:1. Но питание среднестатистического американца далеко и от этого «идеала», и соотношение НЖК в его диете — от 10 до 30:1, что приводит к повышению риска воспалительных заболеваний и других очень серьезных проблем со здоровьем. Такой дисбаланс ставит под угрозу способность тела преобразовывать АЛК в жирные кислоты длинной цепи, такие как ЭПК и ДГК. Согласно данным из разных источников, нам нужно потреблять примерно от 3 до 5% калорий, из ЛК (омега–6) и от 0.5 до 3% калорий из АЛК (омега–3) в день. Всемирная организация здравоохранения рекомендует потребление 5–8% калорий из ЛК и минимум 1% от АЛК, при питании с содержанием жиров, не превышающим 15% от всех калорий. Рекомедованные ВОЗ нормы возрастают при увеличении потребления жиров и наоборот, снижаются при уменьшении жиров в рационе. В 2002 г., Совет по пище и питанию американского Института медицины рекомендовал потребление омега–3 от 1.1 до 1.6 грамм в день. Минимальная рекомендация может всегда использоваться как эффективное средство выявления потребностей в питании. При потреблении 2000 калорий 0.5% калорий из АЛК дает 10 калорий. Это — примерно 1.1 грамма АЛК, или омега–3, что находится в пределах нормы Института медицины (хотя и у нижней границы). Следовательно таким же будет и потребление ЛК, или омега–6, что также укладывается в норму, учитывая минимальное соотношение 1:1. Такое количество НЖК можно без проблем получить, питаясь фруктами и овощами с редкими включениями орехов и семян. Основываясь на вышеуказанных цифрах, при диете 80/10/10 и потреблении 2000 калорий получить оптимальное количество НЖК с пищей можно следующим образом: ● Завтрак: 1.5 фунтов манго (приблизительно 3) и 12 унций черники. ● Второй завтрак: 44 унции бананов (около 11). ● Обед: 1 фунт апельсинов, 1 салата ромэн и 8 унций помидоров. Всего в этих продуктах содержится 1.3 грамма АЛК и 1.4 грамма ЛК. Это соответствует соотношению 1:1 дает необходимый минимум НЖК без употребления какой-либо очевидно жирной пищи. Включение в рацион здорового (очень небольшого) количества очевидных жиров даст гарантию того, что потребности вашего организма удовлетворены. Так как среднестатистический американец потребляет намного больше омега–6, чем омега–3, нам навязывают употребление различных биодобавок, содержащих омега–3. Представление, что мы должны употреблять большое количество отдельно взятого вещества, чтобы уравновесить другое, которое потребляется чрезмерно, столь же неверно, как употребление витамина C, с целью снизить вред от курения сигарет. Увеличение общего потребления жиров, даже «хороших», приведет к тому, что мы будем есть слишком много жира. Мы бы не нуждались ни в каких добавках, если бы мы не вредили себе своим образом жизни и выбором пищи. В нашей идеальной диете продукты уже обладают сбалансированным составом, удовлетворяющим все наши потребности.
14. Фосфолипиды. Строение. Биосинтез фосфолипидов. Фосфолипазы. Фосфолипиды – сложные липиды, одна из спиртовых групп которых связана не с ЖК, а с фосфорной кислотой (фосфорная кислота может быть соединена с дополнительным полярным соединением). Состоят из четырех компонентов, соединенных эфирными связями: 1) спирт 2) жирные кислоты 3) фосфат 4) полярная группировка (чаще всего, серин, холин, этаноламин). В зависимости от содержащегося в них спирта подразделяются на: глицерофосфолипиды (содержат глицерин) и сфингофосфолипиды (содержат сфингозин). В зависимости от содержащегося полярного вещества подразделяются на: фосфатидилсерины, лецитины (фосфатидилхолины), кефалины (фосфатидилэтаноламины). Глицерофосфолипиды Общая формула глицерофосфолипидов имеет следующий вид: Сфингофосфолипиды Общая формула сфингофосфолипидов имеет следующий вид: Сфингозин - это 2-хатомный непредельный аминоспирт. Жирная кислота присоединена пептидной связью к аминогруппе сфингозина. Сфинголипиды делятся на две большие группы: фосфорсодержащие сфинголипиды и гликосфинголипиды. Сфинголипиды широко представлены в живом организме, участвуют в построении мембран, в сложных процессах, связанных с нервной деятельностью животных. Интересно, что содержание фосфорсодержащих сфинголипидов в организме животных увеличивается по мере эволюции его нервной системы: у млекопитающих ах до 10 % от суммы липидов, у рыб — 1—2%. Лецитины (фосфатидилхолины) - сложные эфиры глицерина, в молекуле которых X представлен остатком аминоспирта — холина: Лецитины были впервые обнаружены в желтке яйца, откуда и произошло их название. В фосфолипидах масличных семян и животных содержание фос-фатидилхолинов достигает 30—50%. Кефалины (фосфатидилэтаноламины)— сложные эфиры глицерина, построенные по тому же типу, что и лецитины, но в состав кефалинов вместо холина входит этаноламин: Содержание фосфатидилэтаноламинов в фосфолипидах масличных семян и животных тканях достигает 20—25%. Чаще всего в состав кефалинов входят пальмитиновая, стеариновая и олеиновая кислоты. Кефалины (от греч. голова) впервые были выделены из тканей головного мозга. Фосфатидилсерины - сложные эфиры глицерина, в молекуле которых X представлен остатком аминокислоты — серина: Фосфолипиды - это амфифильные вещества. В молекуле фосфолипидов всегда имеются заместители двух типов: гидрофильные и гидрофобные. Расположение гидрофильных и гидрофобных участков особое. Гидрофильные участки (остаток фосфорной кислоты и полярная группировка) образуют "головку", а гидрофобные радикалы жирных кислот (R1 и R2) образуют "хвосты". Поэтому молекулу фосфолипида обозначают:
Фосфолипиды — бесцветные вещества, без запаха, хорошо растворимы в диэтиловом эфире, хлороформе, плохо в ацетоне. Фосфолипиды являются обязательной составной частью растений и животных; их содержание колеблется в широких пределах. Особенно много их в нервной и мозговой тканях (до 30%). Роль фосфолипидов в жизнедеятельности живого организма чрезвычайно велика. Вместе с белками и другими соединениями они участвуют в построении мембран клеток и субклеточных структур, выполняя роль «несущих конструкций», способствуют переносу химических веществ, а также осуществляют другие функции в биохимических процессах, протекающих в живом организме. В промышленности фосфолипиды получают в качестве побочного продукта при производстве растительных масел. Фосфолипиды нашли широкое применение в медицине и в качестве улучшителей и эмульгаторов в ряде отраслей пищевой промышленности (хлебопекарной, кондитерской, масложировой).
15. Перевариваниелипидов в ЖКТ. Роль желчных кислот и панкреатических липаз. Всасывание липидов в тонком кишечнике, ресинтез липидов в энтероцитах. Гепатоэнтеральная циркуляция желчных кислот.Стеаторея. В полости рта липиды подвергаются лишь механической обработке. В желудке имеется небольшое количество липазы, которая гидролизует жиры. Малая активность липазы желудочного сока связана с кислой реакцией содержимого желудка. Кроме того, липаза может влиять только на эмульгированные жиры, в желудке отсутствуют условия для образования эмульсии жира. Только у детей и у моногастричных животных липаза желудочного сока играет важную роль в переваривании липидов. Кишечник является основным местом переваривания липидов. В двенадцатиперстной кишке на липиды воздействует желчь печени и сок поджелудочной железы, одновременно происходит нейтрализация кишечного содержимого (химуса). Происходит эмульгирование жиров под действием желчных кислот. В состав желчи входят: холевая кислота, дезоксихолевая (3,12 дигидроксихолановая), хенодезоксихолевая (3,7 дигидроксихолановая) кислоты, натриевые соли парных желчных кислот: гликохолевая, гликодезоксихолевая, таурохолевая, тауродезоксихолевая. Они состоят из двух компонентов: холевой и дезоксихолевой кислот, а также глицина и таурина.
Соли желчных кислот хорошо эмульгируют жиры. При этом увеличивается площадь соприкосновения ферментов с жирами и увеличивается действие фермента. Недостаточность синтеза желчных кислот или задержка поступления нарушает эффективность действия ферментов. Жиры, как правило, всасываются после гидролиза, но часть тонко эмульгированных жиров всасывается через стенку кишечника и переходит в лимфу без гидролиза. Эстеразы разрывают в жирах эфирную связь между, спиртовой группой и карбоксильной группой карбоновых кислот и неорганических кислот (липаза, фосфатазы). Под действием липазы жиры гидролизуются на глицерин и высшие жирные кислоты. Активность липазы возрастает под действием желчи, т.е. желчь непосредственно активирует липазу. Кроме того, активность липазы увеличивают ионы Са++ вследствие того, что ионы Са++ образуют нерастворимые соли (мыла) с освободившимися жирными кислотами и предотвращают их подавляющее влияние на активность липазы. Под действием липазы в начале гидролизуются эфирные связи у α и α1 (боковых) углеродных атомов глицерина, затем у β-углеродного атома:
Под действием липазы до 40% триацилглицеридов расщепляются до глицерина и жирных кислот, 50-55% гидролизуется до 2-моноацилглицеринов и 3-10% не гидролизуется и всасываются в виде триацилглицеринов. Стериды корма расщепляются ферментом холестеролэстеразой до холестерина и высших жирных кислот. Фосфатиды гидролизуются под влиянием фосфолипаз А, A2, С и D. Каждый фермент действует на определенную сложноэфирную связь липида. Точки приложения фосфолипаз представлены на схеме: Фосфолипазы поджелудочной железы, тканевые фосфолипазы вырабатываются в виде проферментов и активируются трипсином. Фосфолипаза A2 змеиных ядов катализирует отщепление ненасыщенной жирной кислоты в положении 2 фосфоглицеридов. При этом образуются лизолецитины с гемолитическим действием. фосфотидилхолин лизолецитин Поэтому при попадании этого яда в кровь происходит сильный гемолиз.. В кишечнике эта опасность устраняется действием фосфолипазы A1, быстро инактивирующей лизофосфатид в результате отщепления от него остатка насыщенной жирной кислоты с превращением его в неактивный глицерофосфохолин. Лизолецитины в малых концентрациях стимулируют дифференцировку лимфоидных клеток, активность протеинкиназы С, усиливают клеточную пролиферацию. Коламинфосфатиды и серинфосфатиды расщепляются фосфолипазой А до лизоколаминфосфатидов, лизосеринфосфатидов, которые далее расщепляются фосфолипазой A2. Фосфолипазы С и D гидролизуют связи холина; коламина и серина с фосфорной кислотой и остатка фосфорной кислоты с глицерином. Всасывание липидов происходит в тонком отделе кишечника. Жирные кислоты с длиной цепи менее 10 углеродных атомов всасываются в неэтерифицированной форме. Для всасывания необходимо присутствие эмульгирующих веществ – желчных кислот и желчи. Ресинтез жира, характерного для данного организма, происходит в кишечной стенке. Концентрация липидов в крови в течение 3-5 часов после приема корма высокая. Хиломикроны – мелкие частицы жира, образующиеся после всасывания в кишечной стенке, представляют собой липопротеиды, окруженные фосфолипидами и белковой оболочкой, внутри содержат молекулы жира и желчных кислот. Они поступают в печень, где липиды подвергаются промежуточному обмену, а желчные кислоты проходят в желчный пузырь и далее обратно в кишечник (см. рис.9.3 на стр.192). В результате такого кругооборота теряется малое количество желчных кислот. Считают, что молекула желчной кислоты в сутки совершает 4 кругооборота.
16. Образование и строение липопротеинов. Биологическая роль. Основные пути превращений, роль апопротеинов. Роль липопротеинлипаз в метаболизме липопротеинов (ХМ и ЛПОНП). Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: ТАГ, эфиров холестерина, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки-апопротеины. Компоненты ядра связаны слабыми типами связей и находятся в состоянии постоянной диффузии – способны перемещаться друг относительно друга. Основная роль липопротеинов – транспорт липидов, поэтому обнаружить их можно в биологических жидкостях. При изучении липидов плазмы крови оказалось, что их можно разделить на группы, так как они отличаются друг от друга по соотношению компонентов. У разных липопротеинов наблюдается различное соотношение липидов и белка в составе частицы, поэтому различна и плотность. Липопротеины разделяют по плотности методом ультрацентрифугирования, при этом они не осаждаются, а всплывают (флотируют). Мерой всплывания является константа флотации, обозначаемая Sf (сведберг флотации). В соответствии с этим показателем различают следующие группы липопротеинов:
Липопротеины можно разделить и методом электрофореза. При классическом щелочном электрофорезе разные липопротеины ведут себя по-разному. При помещении липопротеинов в электрическое поле хиломикроны остаются на старте. ЛОНП и ЛПП можно обнаружить во фракции преb-глобулинов, ЛНП - во фракции b-глобулинов, а ЛВП - a-глобулинов:
Липопротеины разделяются соответственно их плотности на четыре основных типа: хиломикроны - ХМ, липопротеины очень низкой плотности - ЛОНП, липопротеины низкой плотности - ЛНП, липопротеины высокой плотности - ЛВП. Существуют также промежуточные формы в метаболизме липопротеинов: хиломикроны остаточные (ХМост), ЛОНП остаточные (или липопротеины средней плотности - ЛСП). Определение липопротеинового спектра плазмы крови применяется в медицине для диагностики атеросклероза. Все эти липопротеины отличаются по своей функции. 1. Хиломикроны (ХМ) - образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном - в жировую ткань), а также - транспорт экзогенного холестерина из кишечника в печень. 2. Липопротеины Очень Низкой Плотности (ЛОНП) - образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань. 3. Липопротеины Низкой Плотности (ЛНП) - образуются в кровеносном русле из ЛОНП через стадию образования Липопротеинов Промежуточной Плотности (ЛПП). Их роль: транспорт эндогенного холестерина в ткани. 4. Липопротеины Высокой Плотности (ЛВП) - образуются в печени, основная роль - транспорт холестерина из тканей в печень, то есть удаление холестерина из тканей, а дальше холестерин выводится с желчью. Транспорт липопротеинов представлен на рисунке 2. Аполипопротеины - это белковая часть липопротеинов (апобелок). В состав липопротеина может входить один или несколько апобелков. Некоторые апобелки являются интегральной частью липопротеина, а другие могут перемещаться с одного липопротеина на другой. Апобелки обозначают буквами: А, В, С, Е. Интегральные апопротеины синтезируются в процессе формирования структуры липопротеина, как, например, белок В-48 в клетках эпителия кишечника. Периферические белки в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя дальнейшие превращения липопротеинов. сунок 2 Например, апопротеин C-II обеспечивает действие фермента липопротеинлипазы и таким образом утилизацию жиров периферическими тканями и превращение хиломикронов в остаточные хиломикроны. Остаточные хиломикроны содержат апопротеин Е, который взаимодействует с рецепторами гепатоцитов, и таким образом остаточные хиломикроны из крови попадают в печень. Функции апопротеинов. 1.Апобелки выполняют функцию эмульгаторов, потому что являются амфифильными веществами. 2.Некоторые из аполипопротеинов являются регуляторами активности ферментов липидного обмена. 3.Могут обладать собственной ферментативной активностью. 4.Могут выступать в качестве лигандов клеточных рецепторов для липопротеинов. 5.Многие апобелки осуществляют транспорт липидов из одного липопротеина в другой. 6.Участвуют в формировании структуры липопротеинов.
17. Ацетил-КоА – как центральный метаболит обмена веществ. Бета - окисление высших жирных кислот в миоцитах как источник энергии. Кетогенез и его биологическое значение. В результате b->:8A;5=8O образуется Ацетил-КоА. В печени образуется больше Ацетил-КоА, чем ей требуется. Печень - "орган-альтруист" и поэтому печень отправляет глюкозу в другие ткани. Печень стремится направить в другие ткани и свой собственный Ацетил-КоА, но не может, так как для Ацетил-КоА клеточные мембраны непроницаемы. Поэтому в печени из Ацетил-КоА синтезируются специальные вещества, которые называются "КЕТОНОВЫЕ ТЕЛА". КЕТОНОВЫЕ ТЕЛА - ЭТО ОСОБАЯ ТРАНСПОРТНАЯ ФОРМА АЦЕТИЛ-КоА! Кетоновые тела синтезируются в печени, легко проходят через митохондриальные и клеточные мембраны и поступают в кровь. Кровью они транспортируются во все другие ткани. Используются только ацетоацетат и бета-гидроксибутират. Их утилизация происходит в митохондриях. Испульзуется в качестве энергии. В норме процессы синтеза и использования кетоновых тел уравновешены, поэтому концентрация кетоновых тел в крови и в тканях обычно очень низка, и составляет 0.12 - 0.30 ммоль/л. Кетогенез-образование кетоновых тел при диабете, голодании, или избыточном употреблении жирной пищи.
18. Распад пуриновых и пиримидиновых нуклеотидов. Особенности и реакции процесса распада, конечные метаболиты. Нарушения обмена пуриновых нуклеотидов при подагре. Начальные этапы этого процесса катализируются специфическими ферментами. Конечные продукты: СО2, NН3, мочевина, β-аланин, β-аминоизомасляная кислота. β-аланин используется для синтеза дипептидов мышц – карнозина и ансерина или выделяется с мочой.
3. Биосинтез пуриновых, пиримидиновых нуклеотидов в тканях. Биосинтез пуриновых мононуклеотидов. Первоначальным соединением синтеза служит Д-рибозо-5-фосфат, который является продуктом пентозофосфатного цикла и на который переносится пирофосфатная группа АТФ. Образовавшийся 5-фосфорибозил-1-пирофосфат (ФРПФ) взаимодействует с глутамином, который является донором NН2-группы в результате чего образуется β-5-фосфорибозил-амин. Эта стадия становится ключевой в синтезе пуринов. Затем присоединяется молекула глицина к свободной NН2-группе β-5-фосфорибозил-амина с образованием глицинамидрибонуклеотида. Еще через несколько стадий образуется первый пуриновый нуклеотид инозинмонофосфат (ИМФ), из которого затем синтезируются остальные нуклеозидфосфаты. Схема:
Биосинтез пиримидиновых нуклеотидов Первоначальными соединениями этого процесса являются карбамоилфосфат и аспарагиновая кислота. Из них через длинную цепь реакций образуется уридинмонофосфат (УМФ) и остальные пиримидиновые нуклеотиды.
19. Классификация сигнальных молекул (микромолекулы, эйкозаноиды, цитокины, гистогормоны, гормоны). Понятие о клетках-мишенях. Основные этапы гормональной регуляции. Пути передачи сигнала клетке (аутокринный, паракринный, эндокринный). Сигнальными молекулами могут быть неполярные и полярные вещества. Неполярные вещества, например стероидные гормоны, проникают в клетку, проходя через липидный бислой. Полярные сигнальные молекулы в клетку не проникают, но связываются специфическими рецепторами клеточных мембран. Такое взаимодействие вызывает цепь последовательных событий в самой мембране и внутри клетки. К полярным сигнальным молекулам относят белковые гормоны (например, глюкагон, инсулин, паратгормон), нейромедиаторы (например, ацетилхолин, глицин, γ-аминомасляная кислота), факторы роста, цитокины, эйкозаноиды. Внутренние - сигналы образуются и действуют в одной и той же клетке, часто сигналами выступают метаболиты. Они выполняются роль аллостерических активаторов/ ингибиторов ферментов. Внешние – управляющие сигналы поступают в клетку из внешней среды. Задачи: Ø Внутреннее и межклеточное согласование метаболических процессов; Ø Исключение холостых циклов метаболизма; Ø Регуляция процессов образования и использования энергии; Ø Поддержание гомеостаза; Ø Приспособление организма к изменениям окружающей среды; Сигнальные молекулы – эндогенные химические соединения, которые в результате взаимодействия с рецепторами обеспечивают внешнее управление биохимическими реакциями в клетках-мишенях. Клетка-мишень - это клетка, имеющая специализированные воспринимающие рецепторы для данного вида сигнальных молекул. Особенности сигнальных молекул: ü Малый период жизни; ü Высокая биологическая активность; ü Уникальность действия; ü Эффект усиления; ü Один вид сигнальной молекулы может иметь несколько клеток-мишеней; ü Реакция разных клеток-мишеней на одну сигнальную молекулу может отличаться; Химические соединения, которые взаимодействуют с определенным рецептором, называют Лигандами. Виды регуляторных эффектов: 1. Эндокринный – сигнальная молекула поступает с током крови к клетке-мишени из желез внутренней секреции (дистантное действие). 2. Паракринный - сигнальная молекула вырабатывается и действует на клетки в пределах одного органа или ткани. 3. Аутокринный - сигнальная молекула действует на клетку её образовавшую. Классификация сигнальных молекул: По химической природе: o Органические – белковые соединения, стероиды и т.д o Неорганические – оксид азота и т.д. По физическим свойствам: o Липофобные - не могут проникать через мембрану клетки. Они растворимы в воде. o Липофильные - растворяются в жирах. Свободно проникают через ЦПМ и действуют на рецепторы внутри клетки. По биологической природе: o Гормоны (по месту образования) сигнальные молекулы с выраженным эндокринным эффектом. o Факторы роста и цитокины – факторы роста. Это сигнальные молекулы белковой природы, которые выделяются неспециализированными клетками организма. Они регулируют рост, дифференцировку, пролиферацию соседних клеток. Действие пара- и аутокринно. o Нейромедиаторы – выделяются нервными клетками и вызывают деполяризацию мембран. сигнальные молекулы, вырабатывающиеся нервными клетками, координирующие работу нейронов и управление периферическими тканями. Их действие связано с влиянием на ионные каналы. Они изменяют их проницаемость и вызывают деполяризацию мембраны. гипоталамус является компонентом и своеобразным «выходным каналом» лимбической системы. Это отдел промежуточного мозга, контролирующий различные параметры гомеостаза. С одной стороны он связан с ЦНС (центры ВНС), с другой - с гипофизом через нервные проводники и особую портальную систему. Гипоталамус участвует во многих функциях нервной регуляции,а также регулирует эндокринную систему. Регуляция метаболизма : внутренняя и внешняя. Внутренняя регуляция - управляющие сигналы образуются и действуют внутри одной и той же клетки (само-регуляция). Внешняя регуляция - управляющие сигналы поступают к клетке из внешней среды. Внутренняя регуляция осуществляется путём изменения активности ферментов активаторами или ингибиторами. Внешняя регуляция обеспечивается специализированными сигнальными молекулами, которые в результате взаимодействия с ферментами обеспечивают внешнее управление биохимическими процессами в клетках-мишенях. Общие этапы действия сигнальных молекул: 1.Распознавание сигналов рецепторами клетки-мишени 2.Передача сигнала и его усиление 3.Изменение биохимических процессов в клетке 4.Элиминация сигнала Вторые посредники в действии липофобных сигнальных молекул, цАМФ и цГМФ -зависимые механизмы действия. Аденилатциклаза, протеинкиназа. Продемонстрировать эффекты гормонов, осуществляющие регуляторное действие при участии цАМФ. Особенности механизма липофобных сигнальных молекул: ü Взаимодействие с поверхностным клеточным рецептором ü Сигнал передается с рецептора внутрь клетки и усиливается там с помощью внутриклеточных регуляторов. Высокомолекулярные вторичные посредники – Мессенжеры, Низкомолекулярные – цАМФ, цГМФ, диацилглицерол, Са. ü Биологическое действие обусловлено сочетанием регуляции активности ранее синтезированных ферментов. Аденилатциклазный МЕХАНИЗМ ДЕЙСТВИЯ, ЗАВИСИМЫЙ ОТ ЦАМФ. Факторы, необходимые для этого: o нерастворимая в воде сигнальная молекула; o поверхностные рецепторы клетки-мишени; o внутриклеточный трансдуктор G-белок. Состоит из 3 единиц: альфа, бета, гамма. · G-белок может быть ингибирующий и активирующий. G-белок способен присоединять ГДФ или ГТФ. o Аденилатциклаза (АЦ) (превращает АТФ в ЦАМФ); o Протеинкиназа ЦАМФ-зависимая. Она катализирует реакцию фосфорилирования белков; o Регуляторные элементы ДНК (ЭЕХАНСЕР и САЙЛЕНСЕР); o ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ; o ФОСФАТАЗА - дефосфорилируют белки; o Белок-синтетический аппарат клетки. Этапы, стимулирующие ЦАМФ -зависимый механизм : 1. взаимодействие сигнальной молекулы с рецептором; 2. изменение конформации G-белка; 3. замена ГДФ на ГТФ в альфа-S единице G-белка;α-субединица отделяется и добавляется к АЦ. 4. альфа-S ГТФ активирует АЦ; 5. АЦ синтезирует ЦАМФ; 6. ЦАМФ активирует ПРОТЕИНКИНАЗУ-А (ПКА); 7. ПКА фосфорилирует белки и белковые факторы транскрипции, изменяющие активность и количество ферментов; 8. Прекращение действия. - отделение α-субединицы от АЦ - ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ. - ФОСФАТАЗА - ДЕФОСФОРИЛИРУЕТ белки. Этапы, ингибирующие ЦАМФ -зависимый механизм: С первого по третий те же самые этапы, отличие в G-белке (альфа-I единица). Четвёртый этап - связывание ГТФ с альфа-I единицей будет ингибировать АЦ. Ингибируюший механизм противодействует и прекращает эффекты ЦАМФ в клетке. ЦГМФ -зависимый стимулирующий механизм действия. цЦГМФ-зависимый механизм Рецептор встроен в мембрану клетки и связан с ферментом ГУАНИЛАТЦИКЛАЗОЙ (ГЦ). При присоединении сигнальной молекулы ГЦ активируется и катализирует реакцию ГТФ * ЦГМФ. Последний активирует ПРОТЕИНКИНАЗУ-G (ПКО), а она запускает реакцию фосфорилирования белков (ферментов и факторов транскрипции). Альдостерон - регуляция объема внутриклеточной жидкости, повышение реабсорбции воды и натрия. Тироксин – повышение основного обмена
20. Эритроциты. Биологическая роль. Гемоглобин: строение, основные этапы синтеза гема, типы и виды гемоглобина. Гемоглобин взрослого организма является тетрамером, состоящим из двух α- и двух β-субьединиц с молекулярными массами примерно 16 кДа. α- и β-цепи отличаются аминокислотной последовательностью, но имеют сходную конформацию. Каждая субъединица несет группу гема с ионом двухвалентного железа в центре. Содержание Hb в крови составляет 140-180 г/л у мужчин и 120-160 г/л у женщин, т. е. вдвое выше по сравнению с белками плазмы (50-80 г/л). Поэтому Hb вносит наибольший вклад в образование рН-буферной емкости крови. Гемоглобин в качестве белкового компонента содержит глобин, а небелкового – гем. Видовые различия гемоглобина обусловлены глобином, в то время как гем одинаков у всех видов гемоглобина. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Атом железа расположен в центре гема-пигмента, придающего крови характерный красный цвет. Каждая из 4 молекул гема «обернута» одной полипептидной цепью. В молекуле гемоглобина взрослого человека HbА содержатся четыре полипептидные цепи, которые вместе составляют белковую часть молекулы – глобин. Две из них, называемые α-цепями, имеют одинаковую первичную структуру и по 141 аминокислотному остатку. Две другие, обозначаемые β-цепями, также идентично построены и содержат по 146 аминокислотных остатков. Таким образом, вся молекула белковой части гемоглобина состоит из 574 аминокислот. Во многих положениях α- и β-цепи содержат разные аминокислотные последовательности, хотя и имеют почти одинаковые пространственные структуры. Получены доказательства, что в структуре гемоглобинов более 20 видов животных 9 аминокислот в последовательности оказались одинаковыми, консервативными (инвариантными), определяющими функции гемоглобинов; некоторые из них находятся вблизи гема, в составе участка связывания с кислородом, другие – в составе неполярной внутренней структуры глобулы. Эритроциты – это высокоспециализированные безъядерные клетки крови. Ядро у них утрачивается в процессе созревания. Эритроциты имеют форму двояковыпуклого диска. В среднем их диаметр около 7,5 мкм, а толщина на периферии 2,5 мкм. Благодаря такой форме увеличивается поверхность эритроцитов для диффузии газов. Кроме того, возрастает их пластичность. За счет высокой пластичности, они деформируются и легко проходят по капиллярам. У старых и патологических эритроцитов пластичность низкая. Поэтому они задерживаются в капиллярах ретикулярной ткани селезенки и разрушаются там. Мембрана эритроцитов и отсутствие ядра обеспечивает их главную функцию – перенос кислорода и участие в переносе углекислого газа. Мембрана эритроцитов непроницаема для катионов, кроме калия, а ее проницаемость для анионов хлора, гидрокарбонат анионов и гидроксил анионов в миллион раз больше. Кроме того, она хорошо пропускает молекулы кислорода и углекислого газа. В мембране содержится до 52% белка. В частности, гликопротеины определяют групповую принадлежность крови и обеспечивают ее отрицательный заряд. В нее встроен Na–К–АТФ–аза, удаляющая из цитоплазмы натрий и закачивающая ионы калия. Основную массу эритроцитов составляет хемопротеин гемоглобин. Кроме того, в цитоплазме содержатся ферменты карбоангидраза, фосфатазы, холинестераза и другие ферменты. Функции эритроцитов: 1. Перенос кислорода от легких к тканям. 2. Участие в транспорте СО2 от тканей к легким. 3. Транспорт воды от тканей к легким, где она выделяется в виде пара. 4. Участие в свертывании крови, выделяя эритроцитарные факторы свертывания. 5. Перенос аминокислот на своей поверхности. 6. Участвуют в регуляции вязкости крови вследствие пластичности. В результате их способности к деформации, вязкость крови в мелких сосудах меньше, чем крупных.
21. Структура гема. Схема распада гемоглобина. Продукты распада гемоглобина в крови, моче, кале. Процесс распада эритроцитов начинается уже в сосудистом русле, а завершается в клеточных элементах системы фагоцитирующих мононуклеаров (купферовских клетках печени, гистиоцитах соединительной ткани, плазматических клетках костного мозга). После выхода гемоглобина из структуры эритроцитов так называемый внеэритроцитарный гемоглобин связывается с гаптоглобином плазмы, образуя комплекс «гемоглобин—гаптоглобин». Благодаря этому гемоглобин задерживается в сосудистом русле, не проходя через почечный фильтр. Вследствие трипсиноподобного действия бета-цепи гаптоглобина и вызванных его влиянием конформационных изменений в порфириновом кольце гема создаются условия для более легкого разрушения гемоглобина в клеточных элементах системы фагоцитирующих мононуклеаров (рис.15). Образующийся таким образом высокомолекулярный пигмент зеленого цвета вердоглобин представляет собой комплекс, состоящий из глобина, разорванной системы порфиринового кольца и трехвалентного железа. Дальнейшие превращения приводят к потере вердоглобином железа и глобина, в результате чего порфириновое кольцо разворачивается в цепь и формируется низкомолекулярный желчный пигмент зеленого цвета —биливердин. Почти весь он ферментативным путем восстанавливается в важнейший красно-желтый пигмент желчи — билирубин, являющийся обычным компонентом плазмы крови. При распаде 1 г гемоглобина образуется 34 мг билирубина. Будучи водонерастворимым, свободный билирубин соединяется с альбумином плазмы, который служит основным его транспортером в плазме крови. Установлено, что каждая молекула альбумина реагирует с двумя (или даже тремя) молекулами билирубина. Комплекс «альбумин—билирубин», доставленный с током крови в печень, на поверхности плазматической мембраны гепатоцита подвергается диссоциации. При этом высвобожденный билирубин образует временный ассоциат с липидами плазматической мембраны и перемещается через нее благодаря деятельности определенных ферментных систем. Дальнейшее прохождение свободного билирубина в клетку происходит при участии в этом процессе двух белков-переносчиков: лигандина (он транспортирует основное количество билирубина) и протеина Z. Лигандин и протеин Z обнаружены также в почках и кишечнике, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детоксикации в этом органе. В клетках печени под влиянием фермента глюкуронилтрансферазы, осуществляющей перенос активных форм молекул глюкуроновой кислоты (уридиндифосфоглюкуроновой кислоты) на свободный билирубин, образуются билирубиндиглюкурониды (до 80%) и билирубинмоноглюкурониды. И те и другие достаточно хорошо растворимы в воде, но лишены способности перемещаться через липидный слой мембраны. За счет связывания билирубина с глюкуроновой кислотой присущая свободному билирубину токсичность в значительной мере теряется. Гидрофобный, липофильный свободный билирубин, легко растворяясь в липидах мембраны и проникая вследствие этого в митохондрии, разобщает в них дыхание и окислительное фосфорилирование, нарушает синтез белка, поток ионов калия через мембрану клеток и органелл. Это отрицательно сказывается на состоянии центральной нервной системы, вызывая у больных ряд характерных неврологических симптомов. В настоящее время под свободным билирубином принято понимать неконъюгированный (с глюкуроновой кислотой) билирубин, который из-за плохой растворимости в воде дает трудную, непрямую (происходящую лишь после внесения в пробу этилового спирта или другого ускорителя, «акселератора») реакцию с ди-азореактивом Ван ден Берга. Поэтому его раньше именовали «непрямой» билирубин. Билирубинглюкурониды (или связанный, конъюгированный билирубин) в отличие от свободного билирубина тотчас вступают в реакцию с диазореактивом («прямой» билирубин). Следует иметь в виду, что в самой плазме крови билирубин, не конъюгированный с глюкуроновой кислотой, может быть либо связан с альбумином, либо нет. Последняя фракция (не связанного ни с альбумином, ни с липидами, ни с другими компонентами крови билирубина) наиболее токсична.
22. Гемолитическая желтуха. Механизм развития. Лабораторные маркеры. Понятие о физиологической желтухе новорожденных. емолитическая желтуха развивается вследствие интенсивного гемолиза эритроцитов при гемолитических анемиях, вызванных сепсисом, лучевой болезнью, переливанием несовместимых групп крови, отравлением сульфаниламидами и т. д. Усиленный гемолиз эритроцитов приводит к интенсивному образованию в клетках РЭС непрямого билирубина. Печень не способна утилизировать в короткое время весь образующийся непрямой билирубин, он накапливается в крови и тканях. Так как печень обезвреживает повышенное количество непрямого билирубина, в больших количествах образуется прямой билирубин в печени. Поступление значительных количеств билирубина в кишечник ведет к усиленному образованию и выделению с калом и мочой стеркобилиногена. Кал приобретает более интенсивное окрашивание. Характерные признаки гемолитической желтухи: · кровь – повышение общего билирубина и непрямого билирубина; концентрация прямого билирубина – в норме; · моча – отсутствие билирубина и положительная реакция на стеркобилиноген (который в больших количествах, чем обычно, поступает в почки из толстого кишечника); · кал – повышение количества стеркобилиногена (темная окраска). Физиологическая желтуха В основе генеза лежат особенности билирубинового обмена у новорожденных, которые проявляются повышенным образованием непрямого биллирубина. (Укороченная жизнь Эр, фетальный Нв, физиологическая полицитемия, несостоятельность эритропоэза, дополнительные источники образования Нв из печеночного цитохрома и миоглобина). Кроме того снижена способность к связыванию и транспортировке непрямого билирубина в крови (мало белка), снижена ф-ция печени, замедленная экскреция коньюгированного билирубина из гепатоцита из-за узости желчных протоков, поступление непрямого билирубина ч/з кишечно-печеночный шунт (минуя печень). Клинически транзиторная гипербилирубинемия проявляется иктеричностью кожи на 2-3 сут, исчезает на 7-8, отсутствие волнообразного течения желтухи. Общее состояние не нарушено, нет гепатолиенального синдрома, билирубин в периферической крови на 3 сут не>205 мкмоль/л, почасовой прирост 5-6 мкмоль/л*час, суточный прирост 85 мкмоль/л*сут, прямой билирубин не>25 мкмоль/л (10-15% от общего).Желтушность проявляется у доношенных при 60 мкмоль, у недоношенных – при 80-100 мкмоль. Особенности билирубинового обмена у новорожденных, обуславливающие высокую частоту желтух: 1. Повышенное образование билирубина (более, чем в 2 раза по сравнению со взрослыми), вследствие: а)укороченной продолжительности жизни эритроцитов (70 - 90 дней у новорожденных и 120 дней у взрослых) из-за преобладания эритроцитов с фетальным гемоглобином, который распадается быстрее) б)повышенным разрушением эритроцитов в костном мозге из-за незрелости эритропоэза в)увеличения образования билирубина их неэритроцитарных источников гема (миоглобина, тканевых цитохромов, предшественников гема) в катаболическую фазу метаболической адаптации 2. Сниженная функциональная способность печени к захвату, конъюгации и экскреции билирубина Повышенная энтерогепатическая циркуляция билирубина, которая обусловлена:а)высокой активностью фермента бета-глюкуронидазы в стенке кишечника б)функционированием в первые дни жизни аранциевого протока, через который часть крови от кишечника поступает в нижнюю полую вену, минуя печень в)стерильностью кишечника. Данные особенности обуславливают высокую частоту неонатальных желтух Поэтому первый вопрос, на который должен ответить неонатолог, курируя ребенка с желтухой: Является желтуха физиологической или патологической? Для Того, чтобы ответить на этот вопрос, нужно знать критерии, отличающие патологическую желтуху от физиологической. (1 признак достаточно) 1.Появление желтухи в первые 24 часа жизни (физиологическая - конец 2 - начало 3 суток) 2.Увеличение уровня билирубина в сыворотке крови более, чем на 85 мкмоль/л в сутки 3.Уровень билирубина на пике подъема более 221 мкмоль/л у доношенных и 257 мкмоль/л у недоношенных 4.Концентрация прямого билирубина более 26 -34 мкмоль/л (10 - 15% от общего количества) 5.Длительность желтухи более 7 - 10 дней у доношенных и более 10 -14 дней у недоношенных.
23. Система гемостаза. Факторы свертывания, строение, место синтеза и биологическая роль. Значение витамина К для синтеза факторов свертывания. Система гемостаза - биологическая система, обеспечивающая, с одной стороны, сохранение жидкого состояния циркулирующей крови, а с другой – предупреждение и купирование кровотечений. Компоненты системы гемостаза: · сосудисто-тромбоцитарное звено · система свертывания крови (коагуляция) · физиологические антикоагулянты · фибринолитическая система (тромболизис) · Фактор I — фибриноген — гликопротеин с молекулярной массой около 340000 дальтон, состоящий из 2946 последовательных аминокислот, и представляет собой димер, в каждой единице которого содержатся три полипептидные цепи, соединенные дисульфидными мостиками. Фактор I в том виде, в каком он вырабатывается паренхиматозными клетками печени и поступает в кровь, называется фибриногеном А, в отличие от фибриногена В, который осаждается из плазмы витамином К (производным b - нафтохинона). Под действием тромбина фибриноген превращается в нерастворимый в крови фибриллярный белок — фибрин, основное вещество (субстрат) тромба (сгустка). В результате увеличения концентрации фибриногена в крови резко повышается вязкость крови, но не усиливается гемокоагуляция. Уменьшением количества фибриногена А ниже 1 г/л иногда обусловлены кровотечения только из-за недостатка фибриногена (гипофибриногенемия). Гипо - и афибриногенемия (полное отсутствие фибриногена в крови) бывают врожденными и приобретенными. Встречается и дисфибриногенемия - состояние, когда под действием тромбина фибриноген крови не превращается в фибрин вследствие функциональной неполноценности молекулы фибриногена. Фибриноген под влиянием тромбина превращается в фибрин по типу протеолитического дробления молекулы фибриногена. Вначале тромбин отщепляет от молекулы фибриногена 2 пептида А, образуя дез-А-мономеры фибрина (неполноценные мономеры фибрина). Затем отщепляются 2 пептида В и возникают дез-АВ-мономеры, или полные мономеры фибрина. Оставшаяся молекула фибриногена — фибрин-мономер. Эта молекула приобретает способность соединяться с себе подобными и образовывать фибрин-полимер, который представляет гель (или сгусток). Сборка мономеров фибрина проходит этапы формирования димеров, из которых при продольном и поперечном сшивании образуются полимеры фибрина — протофибриллы, а затем нити фибрина. Тромб из такого фибрина легко растворяется фибринолизином и потому не может обеспечить полноценный гемостаз. Это нередко бывает причиной кровоточивости и плохого заживления ран. Подобный фибрин называется растворимым (фибрин S, soluble). Полноценным, то есть устойчивым к фибринолизину, он может стать под действием фибриназы (фактора XIIIa). Образовавшийся фибрин называется нерастворимым фибрином (фибрин I, insoluble). · Фактор II— протромбин — относится к эуглобулинам. Под действием протромбиназы образуются a-, b- и g - тромбины. a - тромбин обладает сильной свертывающей активностью в отношении фибриногена, но быстро нейтрализуется антитромбином III. b -тромбин резистентен по отношению к гепарину. g - тромбин не имеет свертывающей активности, но ему присуще фибринолитическое действие. Фактор II синтезируется в печени при участии витамина К. Если нарушается функция печени, концентрация протромбина в крови снижается. Уровень протромбина, или его функция, снижается при эндо- или экзогенной недостаточности витамина К, когда синтезируется неполноценный протромбин. Скорость свертывания крови нарушается лишь при уменьшении концентрации протромбина ниже 40 %. фибринолиз. · Существенной особенностью факторов протромбинового комплекса является зависимость их активности от участия в их синтезе витамина К (рис.1). · Рисунок 1. · При его участии вырабатываемые в печени факторы имеют гамма-карбоксигруппировки, которые осуществляют реакцию с отрицательными группами тромбопластина через кальциевые мостики. Такая реконфигурация неактивного фактора обеспечивает ему раскрытие собственного активного центра и таким образом происходит превращение их в активную форму. · Фактор III — тканевый тромбопластин (неактивная тканевая протромбиназа, апопротеин С—термостабильный липопротеид). Разрушается при 75 °С. Его много в легких, тканях мозга, сердца, кишечника, матки, в эндотелии. Он, в основном, участвует в локальном гемостазе. При контакте с плазменными факторами (VIIa, IV) способен активировать фактор Х (это внешний путь формирования протромбиназы). Из форменных элементов тканевый тромбопластин могут синтезировать только моноциты. · Фактор IV — ионы кальция — имеет первостепенное значение для активации протромбиназы и превращения протромбина в тромбин. Ускоряет фибриноген-фибриновую реакцию. Ионы кальция необходимы для взаимодействия факторов свертывания с фосфолипидной поверхностью клеток. У здоровых людей фактор IV определяется в концентрации 0,8—1,75 ммоль/л. Кальций способен связывать гепарин, благодаря чему свертывание крови ускоряется. Без кальция нарушается агрегация тромбоцитов и ретракция кровяного сгустка. Ионы кальция ингибируют фибринолиз. · Фактор V — проакселерин, лабильный фактор, или Ас-глобулин,— образуется в печени, но, в отличие от других печеночных факторов протромбинового комплекса (II, VII, IX и X), его синтез не зависит от витамина К. Проакцелерин плохо сохраняется в консервированной крови. Он необходим для образования внутренней (кровяной) протромбиназы, при этом заметно активирует фактор X, и для превращения протромбина в тромбин, когда в комплекс включаются фактор Ха, Са2+и фосфолипид. Во время свертывания крови фактор потребляется, как и фактор II, поэтому в сыворотке не обнаруживается. В случаях дефицита фактора V в различной степени нарушаются внешний и внутренний пути образования протромбиназы. Это выражается удлинением протромбинового времени (снижением протромбинового индекса). Тромбиновое время остается в пределах нормы. · Фактор VI — акселерин, или сывороточный Ас-глобулин, - активная форма фактора V. В связи с тем, что отдельным фактором признается только неактивная, профакторная, форма коагулянта, акселерин исключен из употребления и номенклатуры факторов свертывания. · Фактор VII — проконвертин, или конвертин, — синтезируется в печени при участии витамина К. Долго остается в стабилизированной крови, хорошо переносит нагревание, поэтому называется стабильным фактором. Факторы ХII, Ха, калликреин могут превращать фактор VII в VIIa. В основном способствуют образованию тканевой протромбиназы и превращению протромбина в тромбин. Фактор VII в циркулирующей крови активирует фактор X. Это действие усиливается после активации проконвертина тканевым тромбопластином. Врожденным недостатком фактора VII обусловлено развитие геморрагического диатеза. Фактор VII, подобно факторам ХIIа, XI, X, IX, IIа и калликреину, является сериновой протеазой с аргинин-эстеразной активностью. · Фактор VIII — антигемофильный глобулин А, или плазменный тромбопластический фактор А,— относится к сложным гликопротеидам. Место его синтеза точно не установлено. Доказан синтез фактора VIII в печени, селезенке, клетках эндотелия, лейкоцитах, почках. В крови этот фактор циркулирует в виде комплекса из трех субъединиц, обозначаемых VIIIK(коагулирующая единица), VIII-АГ (основной антигенный маркер) и VIII-фВ (фактор Виллебранда, связанный с VIII-АГ). VIII-фВ регулирует синтез коагулянтной части антигемофильного глобулина—VIIIK. При свертывании крови фактор VIII остается в неактивном состоянии. · Фактор IX — Кристмас-фактор, антигемофильный глобулин В, плазменный тромбопластиновый компонент(plasma thromboplastin component— РТС). Фактор IX образуется в печени, поэтому его содержание в крови больных гепатитами, циррозами печени уменьшено. В процессе свертывания крови фактор IX не потребляется и остается в сыворотке еще в более активном состоянии, чем в плазме. Его гемостатический уровень (25 %) достаточен для выполнения хирургических вмешательств. · Фактор Х — фактор Стюарта - Прауэра — гликопротеин с массой молекулы 54200—56000. Вырабатывается в печени в неактивном состоянии при участии витамина К и состоит из двух полипептидных цепей: тяжелой (с молекулярной массой 38000), на которой находится активный центр, и легкой — с остатком карбоксиглютаминовой кислоты, необходимой для присоединения к фосфолипидам. Уровень фактора Х в крови связан с протромбииовым временем. Так, если в крови фактора Х менее 1 %, то протромбиновое время будет более 90 с (при норме 12 - 14 с), если от 1 до 2 % — около 70 - 90 с, если же от 2 до 5 % - 40 - 70 с, а если от 5 до 10 % — 15 - 40 сек. Фактор Х трансформируется в Ха под действием солевых растворов с высокой ионной силой. Для приобретенного или врожденного недостатка фактора Х характерно удлинение протромбинового времени. Вследствие врожденного недостатка фактора X, наследуемого по неполному аутосомному типу, возникает болезнь Стюарта - Прауэра, которая встречается как у мужчин, так и у женщин. Наклонность к кровоточивости определяется только у гомозиготных особей. Содержание фактора Х снижено в крови больных системным амилоидозом, миеломной болезнью, туберкулезом, с поражениями печени, недостатком витамина К, получающих непрямые антикоагулянты. · Фактор XI — РТА (plasma thromboplastin anticedent) — плазменный предшественник тромбопластина - гликопротеин с массой молекулы 160 000. В процессе свертывания крови не потребляется, поэтому обнаруживается в большом количестве в сыворотке. Активная форма этого фактора (ХIа) образуется при участии факторов ХIIа, Флетчера и Фитцджеральда - Фложе. Форма ХIа активирует фактор IX, который превращается в фактор IХа. Эта реакция осуществляется и трипсином в присутствии ионов кальция. Врожденная недостаточность фактора XI наследуется по аутосомно-рецессивному типу. Эта недостаточность выявляется у мужчин и женщин. Кровоточивость в основном отмечается после травм и операций. Фактор XII — фактор контакта Хагемана — соединение с массой молекулы 80 000. Фактор XII вырабатывается в неактивном состоянии. Место его синтеза не известно. В лабораторных условиях активируется при соприкосновении с поверхностью кварца и стекла, каолина, целита, асбеста, углекислого бария; а в организме — при контакте с кожей, волокнами коллагена, хондроитинсерной кислотой, мицеллами насыщенных жирных кислот, бактериальными липополисахаридами, содержащими радикалы жирных кислот, эндотоксином, адреналином и норадреналином. Фактор Хагемана — “инициатор” внутрисосудистой коагуляции, активирует прекалликреины плазмы, которые превращаются в ферменты калликреины, освобождающие кинины, служит активатором фибринолиза. Калликреин активирует фактор XII в 10 раз сильнее, чем плазмин и фактор ХIа. В жидкой среде фактор Флетчера оказывается наиболее важным активатором фактора Хагемана. В крови есть ингибитор активного фактора Хагемана. Врожденный дефицит фактора XII наследуется по аутосомно-рецессивному типу. У больных с недостатком фактора XII в коагулограмме резко удлиняется время свертывания крови. · Фактор XIII — фибринстабилизирующий фактор, фибриназа, фактор Лаки - Лоранда — a2-гликопротеид с массой молекулы 300 000 — 340 000. Определяется в сосудистой стенке, тромбоцитах, эритроцитах, почках, легких, мышцах, плаценте. В плазме находится в виде профермента, соединенного с фибриногеном. Фактор XIII под влиянием тромбина превращается в активную форму (XIIIa). В крови здоровых людей его содержится 80 - 120% (сгусток растворяется за 50—100 с). 10% фибриназы обеспечивают полноценный гемостаз, 2% этого фактора достаточны для остановки кровотечения. Активность фактора XIII хорошо сохраняется при —20 °С. При комнатной температуре активность фермента падает в течение 2—3 дней. В результате прогревания плазмы или сыворотки до 60 °С фибриназа инактивируется за 10 мин. В инкубируемых растворах фибриногена при 40 °С фибриназа разрушается в течение 3 ч. Тромбы, образовавшиеся в присутствии фибриназы, очень медленно подвергаются лизису. Если активность фактора XIII снижается, свертки очень быстро распадаются, даже когда фибринолитическая активность крови нормальная. Установлено, что снижение активности фибриназы сопровождается уменьшением адгезивности и агрегации кровяных пластинок, и, наоборот — при повышении активности фибриназы эти свойства тромбоцитов повышаются. У больных с тромбоэмболическими осложнениями, атеросклерозом, у оперированных, родильниц, после введения адреналина, глюкокортикоидов, питуитрина активность фибриназы повышена, а у больных С-авитаминозом, лучевой болезнью, лейкозом, циррозами, лимфомой, у перенесших адреналэктомию, после приема непрямых антикоагулянтов ее активность снижена.
24. Костная ткань как твердая разновидность соединительной ткани, ее основные функции. Особенности структуры гидроксиапатита и их связь с биологической функцией костной ткани. Костные ткани – специализированный вид соединитель ной ткани с высокой минерализацией межклеточного вещества (костная ткань на 73% состоит из солей кальция и фосфора). Из этих тканей построены кости скелета, выполняющего опорную функцию. Кости защищают головной и спинной мозг (кости черепа и позвоночника) и внутренние органы (рёбра, тазовые кости). Костные ткани состоят из клеток и межклеточного вещества. 25. Гормональная регуляция ремоделирования костной ткани. Роль кальцитриола, паратгормона, кальцитонина, половых гормонов, влияние терапии глюкокортикоидами 26. Нарушения обмена кальция и фосфора при рахите, остеомаляции. Лабораторные маркеры нарушений обмена кальция и фосфора. Организм взрослого человека содержит около 1000 г кальция и 600 г фосфора, 99% кальция и 85% фосфора депонировано в скелете в виде кристаллов гидроксиапатита. В сыворотке крови кальций и фосфат содержатся в связанной с белками и другими соединениями форме, а также в виде ионов. Ионизированные формы кальция и фосфата являются физиологически активными. Содержание общего кальция в сыворотке крови в норме составляет 2,3-2,7 ммоль/л, ионизированного - 1,1-1,4 ммоль/л, неорганического фосфора - 0,7-1,4 ммоль/л. Суточная потребность организма взрослого человека в кальции составляет 0,8-1 г, в фосфоре - 1-1,5 г. На гомеостаз кальция и фосфора оказывают влияние паратиреоидный гормон, кальцитонин и кальцитриол, основными органами-мишенями которых являются кости, почки и кишечник. Паратиреоидный гормон стимулирует освобождение кальция и фосфата из костной ткани и почечный синтез кальцитриола, а также усиливает реабсорбцию кальция и подавляет реабсорбцию фосфата в почках. Кальцитриол активирует резорбцию кости и усиливает почечную реабсорбцию кальция и фосфата и абсорбцию кальция и фосфата в кишечнике. Кальцитонин тормозит остеокластическую резорбцию, тем самым снижая высвобождение кальция и фосфата, а также способствует поступлению фосфата в клетки кости и периостальную жидкость. <pФункции фосфора в организме не сводятся только к формированию минерального компонента кости. Он входит в состав нуклеиновых кислот, фосфопротеинов и фосфолипидов, участвует в метаболизме белков, липидов и углеводов, образовании макроэргических соединений (АТФ, креатинфосфата и др.), поддержании кислотно-основного равновесия, необходим для активации ряда ферментов и нормального функционирования нервов и мышц.</p Ионы кальция принимают участие в мышечном сокращении, выделении нейромедиаторов, контроле возбудимости, внутриклеточном метаболизме, свертывании крови, поддержании целостности мембран, трансмембранном транспорте, высвобождении веществ, синтезируемых в клетке (в том числе гормонов), поступлении в клетку веществ путем фаго- и пиноцитоза, минерализации костей; являются кофакторами многих ферментов и неферментных белков. Кальций воздействует на разнообразные обменные процессы, выполняя функцию внутриклеточного мессенджера (в том числе в комплексе с кальмодулином служит посредником в передаче регуляторных сигналов). Концентрация свободных ионов кальция в цитозоле, поддерживаемая в пределах 0,1-10 мкмоль/л, зависит от активности Са2+-АТФазы, кальциевых каналов и от концентрации кальция во внеклеточной жидкости. Ряд гормонов (а1-адренергические катехоламины, вазопрессин и др.) изменяют проницаемость мембран для кальция, тем самым регулируя его вход в клетку, воздействуя на Nа+/Са2+-обменник. Возможны также как мобилизация кальция из митохондрий и эндоплазматического ретикулума, так и накопление его в этих органеллах. В физиологических условиях происходит постоянное обновление костной ткани - ее резорбция протекает параллельно с образованием остеоида и его минерализацией. При различных нарушениях гормональной регуляции (избыток или недостаток кальцитриола, паратиреоидного гормона, кальцитонин, глюкокортикоидов, эстрогенов и андрогенов, тиреоидных гормонов, инсулина, глюкагона, соматотропина), а также при нарушениях обмена кальция и фосфата сдвигается динамическое равновесие, обеспечивающее стабильное состояние скелета. К заболеваниям с нарушениями метаболизма кальция и фосфора относятся:
27. Витамин Д (холекальциферол). Источники витамина для организма человека. Активные формы витамина, их образование в организме. Участие в обмене кальция и фосфора. Проявления недостаточности (рахит, остеомаляция). Гипервитаминоз Д: возможные причины и проявления. Сначала следует отметить ведущую роль витамина D в минерализации костной ткани. Поскольку с возрастом отмечается физиологическое вымывание кальция, повышается пористость костной ткани. Чтобы предупредить остеопороз, людям старшего возраста рекомендованы витамин D, фосфор и кальций – микроэлементы, образующие неорганический матрикс кости. Больше о процессе минерализации костей и кальций-фосфорном обмене Вы можете почитать здесь. Помимо участия в поддержании прочности костной ткани, витамин D активно выполняет ряд других функций:
Как Вы поняли, дефицит витамина вызывает остеомаляцию, остеопороз, рахит, разрушение зубов, искривление позвоночника, нарушения функций нервов и мышц. Чтобы не допустить недостаточность витамина D, нужно употреблять продукты, где он содержится. При повышенной потребности в нем (беременность, пожилой, детский, подростковый возраст) рекомендованы специальные препараты.
28. Обмен кальция и фосфора. Гипокальциемия, гиперкальциемия: причины и проявления. В раннем детском возрасте (особенно на первом году жизни) заболевания (или состояния), связанные с нарушением фосфорно-кальциевого обмена, занимают ведущее место. Это обусловлено чрезвычайно высокими темпами развития ребенка: за первые 12 месяцев жизни масса тела увеличивается в среднем в 3 раза, длина – в 1,5. Такое интенсивное увеличение размеров тела очень часто сопровождается абсолютным или относительным дефицитом кальция и фосфора в организме. К развитию кальций- и фосфопенических состояний приводят разнообразные факторы: дефицит витаминов (главным образом витамина D), нарушения метаболизма витамина D в связи с незрелостью ряда ферментных систем, снижение абсорбции фосфора и кальция в кишечнике, а также реабсорбции их в почках, нарушения эндокринной системы, регулирующей фосфорно-кальциевый обмен, отклонения в микроэлементном статусе и многое другое. Существенно реже встречаются гиперкальциемические состояния. Они носят, как правило, ятрогенный характер, но представляют не меньшую угрозу организму, чем гипокальциемии. Три узловых момента определяют фосфорно-кальциевый метаболизм в организме: 1. всасывание фосфора и кальция в кишечнике; 2. взаимообмен их между кровью и костной тканью; 3. выделение Ca и P из организма – реабсорбция в почечных канальцах.
Гипокальциемия – это низкий уровень содержания кальция в крови. Медицинская норма составляет 2,2-2,5 моль/литр, когда показатели меньше 1,87, это свидетельствует о наличие заболевания и требует немедленной консультации у специалиста и определенного лечения. Болезнь может носить острый или хронический характер, развиваться у людей всех возрастов, в том числе у детей. К проявлениям относятся парестезии, тетания, а также при тяжелом состоянии — эпилептические припадки, энцефалопатия, сердечная недостаточность. Диагностика основана на определении уровня кальция в плазме. Лечение гипокальциемии включает введение кальция, иногда в комбинации с витамином D. Причины возникновения Гипокальциемию могут вызывать заболевания, приводящие к нарушению освобождения кальция из костей или хронической потере кальция вместе с мочой. Поскольку большая часть кальция связана с альбумином, то недостаток данного белка вызывает снижение уровня кальция в крови. Но вызванная недостатком альбумина гипокальциемия не приводит к каким-либо последствиям, так как симптомы гипокальциемии определяются лишь содержанием кальция, не связанного с альбумином. Причинами гипокальциемии также выступают: · различные новообразования; · болезни паращитовидных желез или их резекция; · токсическое поражение печени при отравлении некачественным алкоголем или солями тяжелых металлов; · гипоальбуминемия; · дефицит витамина D; · недостаточное поступление кальция с пищей при голодании, исключение из рациона молочных продуктов, · обильное потоотделение; · беременность и кормление ребенка грудным молоком; · нарушение процесса всасывания кальция по причине диареи, резекции кишки, недостаточной функции поджелудочной железы; · гиподинамия; · аллергические реакции; · гипернатриемия; · прием кортикостероидов и интерлейкинов; · прием препаратов для лечения гиперкальциемии: антиконвульсантов (фенитоина, фенобарбитала) и рифампина; · применение радиоконтрастных агентов, которые содержат этилендиаминтетраацетат; · переливание цитратной крови (более 10 единиц); Причинами гипокальциемии могут быть шоковое состояние или сепсис. проявлению симптомов гипокальциемии ведут гипопаратиреоз, снижение функций паращитовидных желез, которые отвечают за процесс обмена кальция в организме. Еще одной причиной гипокальциемии выступает псевдогипопаратиреоз, который представляет собой наследственное заболевание, связанное с нечувствительностью организма к паратиреоидному гормону, что вызывает снижение уровня кальция и повышение концентрации фосфора. Снижению уровня кальция могут способствовать болезнь Олбрайта, почечная недостаточность, сопровождаемая нарушением обмена витамина D, фосфора и солей кальция. При наличии злокачественных новообразований, гиперкальциемия может возникнуть вследствие метастаз в кости опухоли, а также ввиду усиленной продукции опухолевых клеток, провоцирующих резорбцию в костной ткани. Кроме того, данное заболевание может возникнуть также по причине синтезируемого опухолевыми клетками паратиреоидного гормона и под воздействием иных специфических причин. Гиперкальциемия провоцирует образование спазма афферентных артериол, также она снижает уровень почечного кровотока. При заболевании снижается клубочковая фильтрация, происходящая в отдельно рассматриваемом нефроне и в целом в почке, угнетению подвергается реабсорбция калия, магния и натрия в канальцах при одновременном повышении реабсорбции бикарбоната. Также важно отметить и то, что при этом заболевании увеличивается экскреция (выведение из организма) ионов водорода и кальция. За счет сопутствующего нарушения в функциях работы почек объясняется значительная часть тех проявлений, которые в целом присущи гиперкальциемии. ГИПЕРКАЛЬЦИЕМИЯ: СИМПТОМЫ Ранние симптомы заболевания проявляются в таких состояниях: Потеря аппетита; Запор; Тошнота; Рвота; Боли в животе; Избыточная выработка почками мочи (полиурия); Частый вывод жидкости из организма, приводящий к обезвоживанию со свойственными ему симптомами. В острой форме проявлений гиперкальциемия характеризуется следующими симптомами: Функциональные нарушения работы мозга (эмоциональные расстройства, спутанность сознания, галлюцинации, бред, кома); Слабость; Полиурия; Тошнота, рвота; Повышение давления при дальнейшей его смене развивающейся дегидратацией гипотонии и последующим коллапсом; Заторможенность, ступор. Хроническая гиперкальциемия характеризуется меньшей выраженностью неврологической симптоматики. Возможным становится формирование почечных камней (с кальцием в их составе). Полиурия, наряду с полидипсией, развивается за счет снижения концентрационных с нижения концентрационных способностей почек по причине нарушений в активной транспортировке натрия. Ввиду уменьшения объемов внеклеточной жидкости усилению подвергается реабсорбция бикарбоната, что оказывает способствующее влияние на развитие метаболического алкалоза, повышение же экскреции калия и секреции приводит к гипокалиемии. При тяжелой и длительной гиперкальциемии почки претерпевают течение в них процессов с образованием кристаллов с кальцием, вызывающих серьезные поражения необратимого масштаба. ГИПЕРКАЛЬЦИЕМИЯ: ПРИЧИНЫ ЗАБОЛЕВАНИЯ Развитие гиперкальциемии может быть спровоцировано за счет повышения уровня всасываемости в ЖКТ кальция, а также при избытке его поступления в организм. Нередко наблюдается развитие заболевания среди людей, принимающих значительное количество кальция (к примеру, в процессе развития у них язвенной болезни) и антацидов, также содержащих в себе кальций. В качестве дополняющего фактора выступает употребление больших объемов молока в рационе питания. Оказывает собственное влияние на повышение концентрации в крови кальция и переизбыток витамина D, который, кроме того, способствует увеличению его всасывания через ЖКТ. Между тем, чаще всего гиперкальциемия возникает из-за гиперпаратиреоза (избыточной выработки паратгормона одной или несколькими паращитовидными железами). Порядка 90% из общего числа больных с диагнозом первичного гиперпаратиреоза сталкиваются с обнаружением у них доброкачественной опухоли одной из указанных желез. Для 10% остальных актуальным становится обыкновенное увеличение при выработке гормона в избыточном количестве. Крайне редким, однако не исключаемым явлением становится образование злокачественных опухолей паращитовидных желез за счет гиперпаратиреоза. Клинические проявления гиперкальциемии Преимущественно гиперпаратиреоз развивается среди женщин и пожилых людей, а также среди тех пациентов, которые прошли через лучевую терапию шейной области. В некоторых случаях гиперпаратиреоз образуется в качестве такого редкого наследственного заболевания, как множественная эндокринная неоплазия. Достаточно частым явлением становится гиперкальциемия для больных с имеющимися у них злокачественными опухолями. Так, злокачественные опухоли, локализующиеся в легких, яичниках или почках начинают вырабатывать белок в избыточном количестве, он же впоследствии воздействует на организм по аналогии с паратгормоном. В конечном итоге это образует паранеопластический синдром. Распространение (метастазирование) злокачественной опухоли возможно к костям, чему сопутствует уничтожение костных клеток при одновременном их способствовании к выделению в кровь кальция. Данное течение свойственно опухолям, образующимся в частности в области легкого, молочной и предстательной желез. Поражающая костный мозг злокачественная опухоль также может способствовать разрушению кости наряду с гиперкальциемией. В процессе развития иного типа злокачественных опухолей повышение концентрации в крови кальция на данный момент объяснениям не поддается по причине неполной изученности подобного течения патологии. Примечательно, что гиперкальциемия также может являться спутником многих заболеваний, при которых происходит разрушение костей либо потеря кальция. В качестве одного из подобных примеров можно выделить болезнь Педжета. Развитию гиперкальциемии может способствовать и нарушение подвижности, что в частности актуально при параличах или длительности пребывания в постельном режиме. Эти состояния также приводят к потерям кальция костной тканью при последующем его переходе в кровь. 29. Поступление в организм кальция и фосфатов, пути выведения, биологическая роль. Кальций является структурным макроэлементом, содержание которого превышает по своему содержанию все остальные элементы в организме (кроме элементов-органогенов). Биологическая роль кальция Прежде всего, кальций является важнейшим структурным компонентом костей и зубов.
30. Химический состав эмали зубов. Теоретическая модель структуры эмали. Минерализация эмали в период формирования зубов. Неорганические вещества зрелой эмали составляют 94-95%, в незрелой формирующейся эмали их намного меньше - всего 5%, а в эмали молочных зубов - 80%. После удаления минеральных компонентов остается тонкая сеть органической матрицы. Кроме солей фосфата кальция в составе эмали обнаружены свыше 30 разных элементов. В относительно больших количествах при- сутствуют ионы Mg2+, Na+, а также Cl-, K-, Zn2+и Fe2+. Минеральный состав эмали может колебаться в зависимости от характера питания, но процентное соотношение кальция, фосфора и карбоната довольно постоянно. Содержание Sr2+, Pb2+ и некоторых других микроэлементов в эмали колеблется значительно и зависит от их количества в почве данной местности. Минеральные вещества в эмали распределены неравномерно. Поверхностные более плотные слои содержат меньше воды, карбонатов и больше фтора. Количество неорганических компонентов уменьшается в направлении от поверхности к зоне перехода эмали в дентин (т аб л. 4.1). Содержание кальция и фосфора в эмали соответственно составляет 33,6-39,4 и 16,1-18,0% по отношению к остальным элементам эмали и в направлении от поверхности зуба к дентину их содержание снижается. Обычно снаружи она для ионов Ca2+ составляет 37,8, а внутри - 34,5% и для фосфатов - 18 и 15%. Однако при этом соотно- Таблица 4.1 шение кальция и фосфатов остаётся постоянным (2,1 и 2,3 - весовое и 1,62-1,78 - молярное соотношение). Такая же закономерность распределения концентрационного градиента в эмали относится и к хлоридам. Напротив, содержание карбонатов, натрия, магния и железа в эмали увеличивается по направлению к дентину. Свинец присутствует в низких концентрациях. Он накапливается в поверхностных слоях эмали, в то время как медь и стронций равномерно распределяются по всей толщине эмали. Этапы минерализации: Первичная минерализация эмали представляет двухступенчатый процесс, включающий инициацию и последующий рост кристаллов (эпитаксию). Вначале формируются длинные и тонкие кристаллиты, которые встраиваются в органический матрикс параллельно друг другу. В более позднем периоде кристаллиты утолщаются и превращаются в плоские шестиугольные призмы. Упорядоченное построение и форма кристаллов эмали отличается от бесформенных пластинчатых призм кристаллов кости и дентина. Рост кристаллов регулируется ионами Ca2+ и PO43. В свою очередь, поток жидкости, изменяющийся в течение развития эмали, регулирует эмалевый матрикс. В регуляции роста кристалла в длину, ширину и толщину участвуют амелогенины, упакованные в наносферы. Вторичная минерализация эмали Созревание эмали сопровождается значительным снижением содержания органических компонентов. Происходит распад амелогенинов и задерживается деградация энамелинов, при этом энамелины прочно связываются с кристаллами апатита. Образованная первичная эмаль является незрелой. Она состоит на 30% из органического матрикса и на 70% - из минеральных солей. Во вторичной минерализации участвуют энамелобласты стадии созревания, которые содержат большое количество кальций-связывающих белков. Через энамелобласты к эмали переносятся неорганические ионы и удаляются из созревающей эмали органические вещества и вода. Третичная минерализация эмали Окончательная минерализация эмали происходит уже после прорезывания зуба, и особенно интенсивно - в течение первого года нахождения коронки зуба в полости рта. Часть неорганических веществ поступает со стороны дентина, но основное их количество поставляет слюна. В связи с этим для полноценной третичной минерализации очень важен минеральный состав и рН слюны. Сформированная эмаль лишена способности к росту и не способна к регенерации. После прорезывания зубов продолжается процесс минерализации - "созревание " эмали. С возрастом происходит снижение ее проницаемости. Эмаль является тканью с очень низкой интенсивностью обменных процессов.
31. Кариес. Современные подходы к профилактике и лечению кариеса. Понятие о реминерализации эмали. Кариес зубов – это разрушительный процесс дистрофического или инфекционного характера, протекающий в кости или надкостнице и заканчивающийся полным или частичным разрушением зуба. Причины развития Кариозная болезнь является самым распространенным заболеванием среди взрослого населения. По данным ВОЗ заболеваемость кариесом зубов в разных странах и среди разного контингента колеблется от 80% до 98%. В последние два десятилетия отмечается тенденция роста заболеваемости среди детей, особенно в экономически развитых странах и уже к 6-7 годам у 80-90% детей наблюдается кариес разной глубины. Кариозная болезнь не является самостоятельным заболеванием и важным патогенетическим звеном в ее развитии выступают патологические состояния организма в целом. Так, снижение общего и местного иммунитета, патологии желудочно-кишечного тракта и погрешности в диете способствуют развитию кариеса. На сегодняшний день теорий о возникновении кариеса около четырехсот, но большая их часть основана на том, что из-за нарушения гигиены полости рта на эмали появляется зубной налет, который и приводит к развитию кариозной болезни. Налет появляется из-за неправильной и нерегулярной чистки зубов, особенно в труднодоступных для чистки местах и в местах, откуда он не удаляется естественным путем во время жевания (боковые поверхности зубов, углубления жевательных поверхностей). Зубной налет прочно связывается с поверхностью зуба и является средой обитания для бактерий, большую часть из которых составляют стрептококковая флора. Минеральные соли, которые содержатся в слюне, способствуют уплотнению налета. Такие образования из зубного налета и минеральных солей называют зубной бляшкой. Бактерии, обитающие в зубной бляшке, продуцируют молочную кислоту, которая деминерализует зубную эмаль. Деминерализация зубной эмали – это первый этап кариозного процесса. Полисахарид декстран, который продуцируется стрептококками из сахарозы, способствует процессу деминерализации, именно поэтому развитие кариеса связывают с употреблением в пищу большого количества простых углеводов. Активность микроорганизмов в зубной бляшке и процесса деминерализации зависит от индивидуальных особенностей организма. У большинства людей резистентность к кариесогенным бактериям выражена слабо, тогда как у людей с хорошим состоянием иммунной системы и при отсутствии сопутствующих заболеваний резистентность достаточно высока. У лиц, которые имеют иммунодефицитные состояния, кариес развивается более активно. А у детей с экссудативным диатезом и рахитом кариозная болезнь диагностируется в 2 раза чаще. При изменении состава слюны, когда в ней нарушается соотношение минеральных солей и снижаются ее естественные антибактериальные свойства, увеличивается риск развития кариеса. Соматические заболевания и погрешности в диете с дефицитом минералов, особенно в период формирования зубов, существенно снижают резистентность. Наследственные патологии эмали (аплазия или гипоплазия эмали) и экстремальные воздействия на организм в сочетании с другими факторами повышают вероятность возникновения кариозной болезни. Проживание в промышленных районах, неблагоприятные экологические условия и неполноценность питьевой воды снижают общие защитные функции организма, что может стать важным патогенетическим звеном в появлении кариеса. Липкие остатки углеводистой пищи и отклонения биохимического состава твердых тканей зуба являются основными местными факторами, которые способствуют развитию кариозного процесса. Состояние зубочелюстной системы в период закладки, развития прорезывания и формирования зубов имеет большое значение в дальнейшем состоянии зубочелюстной системы. Клинические проявления В зависимости от глубины поражения твердых тканей зуба выделяют 4 формы кариеса. На стадии кариозного пятна происходит помутнение эмали зуба. Видимых разрушений твердых тканей зуба нет, инструментальное обследование зондом малоинформативно, так как на этом этапе еще нет признаков изменения структуры твердых тканей зуба. Иногда возможен регресс пятна, причины этого явления не выяснены, однако стоматологи связывают самовыздоровление с активизацией иммунной системы. Вторая форма кариеса – это поверхностный кариес. На поверхности зуба появляется темная пигментация, во время инструментального осмотра в зоне пигментации выявляется размягчение эмали. Иногда уже на стадии поверхностного кариеса в разрушительный процесс вовлекаются все слои зубной эмали. Но обычно дефект ограничен и не выходит за пределы эмали. Кариозное поражение выглядит как грязно-серое или коричневое пятно с шероховатым дном. При среднем кариесе поражаются ткани эмали и дентина, а при глубоком кариесе происходит полное поражение всех тканей зуба, вплоть до полного разрушения зуба. Субъективные ощущения зависят от глубины поражения и остроты течения кариеса. Обычно пациенты жалуются на острую боль при попадании на пораженную поверхность или в кариозную полость кислой, сладкой или холодной пищи. При устранении раздражающего фактора острая зубная боль затихает, как правило, самопроизвольных болей при кариесе не бывает. При остром течении кариеса одновременно поражается ряд зубов, пораженные ткани грязно-серого цвета, размягчены, очаг разрушения имеет неправильные очертания, его края подрыты, болевой синдром выражен ярче. Острейшее течение кариеса характеризуется поражением почти всех зубов, при этом в каждом зубе имеется несколько очагов кариозного поражения. Хроническое течение кариозной болезни характеризуется пигментацией пораженных участков, их уплотнением и сглаженностью краев. Поражены отдельные зубы, обычно процесс имеет вялотекущее течение. При отсутствии лечения кариес осложняется пульпитом и периодонтитом, что является основной причиной разрушения зубов и их последующего удаления. Диагностика и лечение Диагностируют кариес во время визуального и инструментального осмотров стоматолога. Субъективные ощущения пациента позволяют судить о глубине процесса, но не являются основными диагностическими критериями. Основным принципом лечения кариеса является удаление пораженных тканей и восстановление зуба пломбировочными материалами. Во время санации кариозной полости удаляются все пораженные ткани. Полость тщательно дезинфицируется, иногда для этого устанавливают временные пломбы, под которыми находятся дезинфицирующие средства. Чем лучше произведена дезинфекция кариозной полости, тем прочнее будет держаться пломба. Качественное укрепление ослабленных тканей зависит как от качественного удаления кариозного очага и его дезинфекции, так и от того, на каком этапе развития кариозной болезни начато лечение. Классическое препарирование кариозного очага происходит с помощью бормашины, однако сегодня имеется альтернативный вариант – лазерное препарирование. Процедура отличается безболезненностью, бесшумностью и качеством подготовки полости зуба к дальнейшему пломбированию. Лечение поверхностного кариеса возможно несколькими способами. Распространенным методом является препарирование зуба и последующее пломбирование кариозной полости. Обязательным условием для этого метода лечения является создание полости в пределах дентина, но при создании мелкой полости условий для создания двухслойной пломбы из изолирующей прокладки и собственно пломбы не достаточно. Эти факторы учитываются при пломбировании кариозных полостей на жевательных и контактных поверхностях премоляров и моляров. Другая методика заключается в применении новых композиционных материалов с высокими агдезивными свойствами, что позволят пломбировать поверхностный кариес без глубокого препарирования тканей зуба. Третий метод заключается в сошлифовывании кариозного участка с последующей реминерализацией. Реминерализацию эмали проводят с помощью аппликаций или электрофореза с 1% раствором фторида натрия (глубокое фторирование зубов) либо с помощью других разрешенных реминерализующих препаратов. При среднем кариесе возможен только метод препарирования твердых тканей зуба с последующим пломбированием кариозной полости. Лечение глубокого кариеса сопряжено с определенными сложностями, так как восстановление почти полностью разрушенного зуба требует от врача не только профессионализма, но и выбора методики. Иногда при глубоком кариесе пульпа зуба покрыта слоем неповрежденного дентина, в некоторых случаях врач вынужден оставлять в кариозной полости пигментированный и размягченный дентин. Такие случаи глубокого кариеса требуют наложения лечебной прокладки на дно кариозной полости (чаще всего используется кальцемин-паста). Эта прокладка оказывает противовоспалительное действие и стимулирует дентиногенез. То есть пломба при глубоких кариозных поражениях состоит из трех слоев: лечебная прокладка, изолирующая фосфатноцементная прокладка и постоянный пломбировочный материал, чаще всего это амальгама. Выбор пломбировочного материала основан на групповой принадлежности зубов. Для того, чтобы в результате пломбирования восстановить анатомические формы передних зубов – резцов и клыков, к пломбе предъявляются требования, как по прочности, так и по эстетичности. Для пломбирования этих групп зубов материал выбирается в соответствии с цветом зубов пациента, что позволяют силикатные цементы и композиционные материалы. Обязательно соблюдение технологического процесса при приготовлении пломбировочного материала, потому как использование хороших компонентов при несоблюдении технологии приготовления материала и при несоблюдении техники установки пломбы существенно снижает ее долговечность. Подготовка кариозной полости к пломбированию заключается в тщательном удалении дентинных опилок с помощью струи воды или воздуха. Далее полость дезинфицируют и высушивают, потому как даже незначительные следы влаги существенно ухудшают прилипаемость цементов, светополимеров и композитных пломб. После затвердевания пломбы ее шлифуют и полируют, удалив перед этим излишние выступы. Чем лучше отшлифована поверхность пломбы, тем меньше на ее поверхности задерживаются микроорганизмы и остатки пищи. Однородность наружного слоя снижает вероятность коррозии пломбы и предотвращает ее быстрое разрушение. Лечение даже хронического и запущенного кариеса с помощью пломбирования позволяет восстановить целостность поверхности зуба, предотвратить пульпит и полное разрушение зубов. Профилактикой кариеса является борьба с мягким зубным налетом, что включает использование качественных зубных паст и зубных щеток, использование зубной нити, регулярное прохождение процедуры профессиональной гигиены полости рта. Если зубная эмаль ослаблена, то для ее укрепления показана флюоризация зубов с помощью фторсодержащих препаратов: паст, растворов и лаков. Сбалансированное питание с высоким содержанием твердой пищи и низким содержанием простых углеводов предотвращает размножение микроорганизмов в полости рта. В районах, где качество питьевой воды низкое, рекомендуется употреблять воду из других регионов.
32. Кариес. Теоретические основы патогенеза и профилактики. Стадии формирования зубного налета и его кариесогенность. Кариес зубов - патологический процесс, многофакторный, проявляющийся после прорезывания зубов, при котором происходит потеря твердыми тканями (эмалью, дентином и/или цементом) минеральных веществ (деминерализация) и их размягчение с последующим образованием полости.
В основе патогенеза кариеса лежит разрушение структуры ГА эмали вследствие усиления деминерализации. В чем причина усиления деминерализации? Первопричиной усиления деминерализации является повышение концентрации протонов в среде, окружающей зуб, снижение рН. Снижение рН приводит к следующим последствиям. 1. Изменяется ионный обмен в ГА эмали, в кристаллическую решетку включается большое количество Н+. Вследствие включения Н+ повышается растворимость эмали, увеличивается выход прежде всего ионов кальция. Если, происходит снижение соотношения Са/Р ниже 1,3 кристаллическая решетка ГА разрушается. 2. Сдвигается равновесие в системе кристалл ГА - раствор ГА (раствор ионов Са2+ и НРО42-). Произведение молярных концентраций ионов кальция и гидрофосфата становится ниже ПР и не обеспечивает нужной скорости минерализации эмали. Таким образом, при снижении рН происходит снижение скорости минерализации и увеличение скорости деминерализации. Говорят "идет кислотное растворение эмали". Кислотное растворение при кариесе происходит локально в области контакта с эмалью мягкого зубного налета. Методы очистки белков Наиболее трудоёмкий этап получения индивидуальных белков - их очистка от других белков, находящихся в растворе, полученном из данной ткани. Часто изучаемый белок присутствует в небольших количествах, составляющих доли процента от всех белков раствора. Так как белки обладают конформационной лабильностью, при работе с белками следует избегать денатурирующих воздействий, поэтому выделение и очистка белков происходят при низких температурах. На первых стадиях очистки белков целесообразно использовать методы, учитывающие какую-либо характерную особенность данного белка, например термостабильность или устойчивость в кислых растворах. Первыми методами очистки необходимо удалить из раствора основную массу балластных белков, которые значительно отличаются от выделяемого белка физико-химическими свойствами. Впоследствии применяют всё более тонкие методы очистки белка. Очистка белков избирательной денатурацией Большинство белков денатурирует и выпадает в осадок уже при кратковременном нагревании раствора до 50-70°С или подкислении раствора до рН 5. Если выделяемый белок выдерживает эти условия, то с помощью избирательной денатурации можно удалить большую часть посторонних белков, отфильтровав выпавшие в осадок-белки, или осадить их центрифугированием. Высаливание Метод очистки белков, основанный на различиях в их растворимости при разной концентрации соли в растворе. Соли щелочных и щёлочно-земельных металлов вызывают обратимое осаждение белков, т.е. после их удаления белки вновь приобретают способность растворяться, сохраняя при этом свои нативные свойства. Чаще всего для разделения белков методом высаливания используют разные концентрации солей сульфата аммония - (NH4 )2 SO4 . Чем выше растворимость белка, тем большая концентрация соли необходима для его высаливания. |
Последнее изменение этой страницы: 2019-03-21; Просмотров: 71; Нарушение авторского права страницы

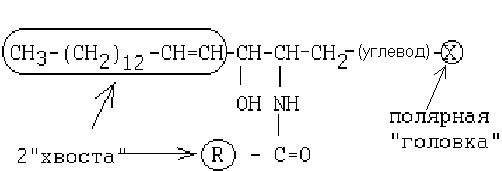
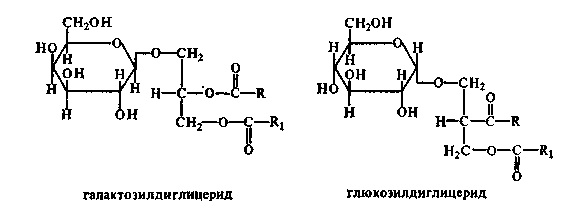 Гликолипиды классифицируют в зависимости от строения углеводного компонента на две группы.
Гликолипиды классифицируют в зависимости от строения углеводного компонента на две группы.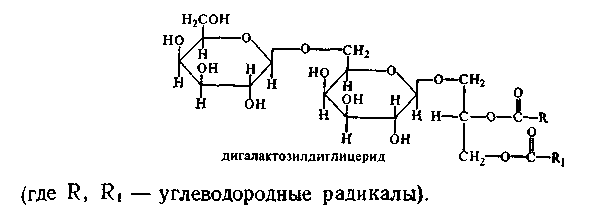 Мыла и детергенты
Мыла и детергенты